

Mirando ao ollo
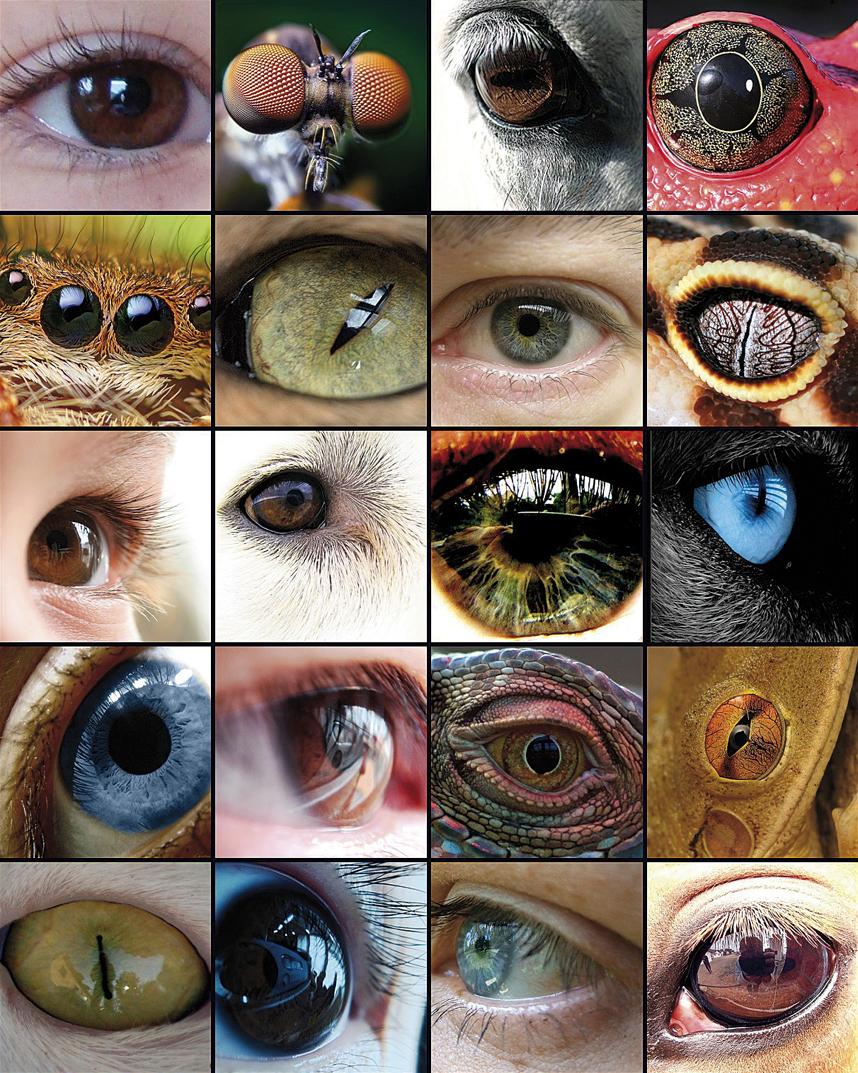
A evolución do ollo foi a base de numerosos estudos, xa que é un exemplo representativo do órgano homólogo nos diferentes taxones. Aínda que algúns compoñentes do ollo, como os pigmentos visuais, parecen ter un precursor común, os ollos capaces de producir imaxes complexas evolucionaron entre 50 e 100 veces, recorrendo ás mesmas proteínas e xenes de forma recorrente.
O propio Charles Darwin recoñecía no seu libro A orixe das especies (1882) que tiña grandes dificultades paira explicar a evolución do ollo e dedicou un capítulo completo ao ollo baixo o título As dificultades da teoría dicindo: "o feito de que un órgano tan complexo como o ollo sexa creado por elección parece, certamente, una tolemia enorme". Pero logo seguía así: "Por iso, si dun ollo simple e imperfecto producíronse os cambios necesarios paira crear o ollo perfecto e complexo de hoxe, tal e como se produciron en realidade, e si aos animais portadores deses cambios resultábanlles útiles ante a contorna cambiante da vida, o feito de que os nosos ollos complexos perfectos formásense por selección natural, aínda que supere a nosa imaxinación, non parece una idea tan revolucionaria".
Orixe das células fotorreceptoras nos metazoos
As células fotorreceptoras son células nerviosas da retina capaces de levar a cabo a fototransducción. Os fotorreceptores son de gran importancia biolóxica mediante a absorción de fotóns e a realización de numerosos carreiros bioquímicos complexos que converten o sinal recibido en imaxe no cerebro.
Existen dúas hipótese paira explicar a orixe das células fotorreceptoras dos metazoos: a primeira baséase na diferenciación e a segunda na simbiose. Segundo a primeira hipótese, os metazoos formáronse a partir de una colonia de flagelados, e todas as células tiñan desde o principio un orgáculo fotorreceptor. Este orgánulos fotorreceptor actuaba como cicatriz ocular e como resposta á fototaxis que transmitía os sinais a través do flagelo.

Segundo a hipótese da simbiose, teñen a súa orixe en flagelados como Volvox ou Chlamydomonas, nos que o orgáculo fotosensible atópase en cloroplastos, o que suxire que a percepción da luz atópase nunha evolución a partir das cianobacterias e que posteriormente se integra como cloroplasto en células eucariotas. Outra proba a favor desta hipótese, explicou Greuet en 1965, ao describir os orgánulos fotorreceptores de dinoflagelados como Erythropsis e Warnovia, tan desenvolvidos como o ollo humano pero agregados no interior dunha soa célula, cunha superficie similar á córnea, una estrutura similar ás lentes, outra similar á retina e orgánulos pigmentos. Os dinoflagelados, debido aos simbiontes comúns de corais, anemones mariñas e outros cnidarios, podían transferir aos cnidarios os xenes necesarios paira responder á luz, o que explicaría a aparición súbita do ollo aos cnidarios.
Segundo a hipótese dos Sinbiontes, a sensibilidade á luz xurdiu primeiro nas cianobacterias. Estas cianobacterias foron interiorizadas por células eucariotas de algas vermellas como cloroplastos primarios. Posteriormente, as algas vermellas pasaron a estar en simbioses con dinoflagelados como cloroplastos secundarios. Nalgunhas especies de dinoflagelados, como Erythropsis e Warnovia, non se atoparon cloroplastos secundarios que se cre que evolucionaron e convertéronse en orgánulos fotorreceptores efectivos, como suxire Greuet. Por último, e dado que os dinoflagelados son sinbiontes comúns aos cnidarios, transmitiron a estes os xenes dos fotorreceptores. Este último é o paso máis incerto do modelo, aínda que se atoparon varios xenes dos dinoflagelados integrados no xenoma dos cnidarios.
Análise do desenvolvemento do ollo
Nos animais é fácil detectar mutacións que afectan o desenvolvemento do ollo e a mutación eyeless (ey) na mosca Drosophila foi descuberta por primeira vez en 1915 polo investigador Hoge. No rato atopouse una mutación parecida e denominouse small eye porque os animais heterocigóticos tiñan os ollos moi reducidos, mentres que os fetos de homocigótanos que morrían no útero faltaban os ollos, pero tamén o nariz e algunhas partes do cerebro (Hill et ao ., 1991).
No home, unha síndrome hereditaria chamado aniridia produce un fenotipo moi parecido. Os xenes Small eye e aniridia foron clonados por Walther e Gruss (1991) e Ton e o seu grupo (1991) respectivamente, e viron que ambos respondían o xene Pax6 extremadamente conservado. Quiring e o seu equipo (1994) clonaron ao homólogo Pax6 de Drosophila e viron sorprendido que o eyeless (ey) de Hoge era igual ao xene. A existencia de mutacións de xenes homólogos Small eye, aniridia e eyeless indicaba que o xene Pax6 era o principal xene de control do desenvolvemento ocular tanto en vertebrados como invertebrados.
Paira demostrar esta hipótese, o investigador WJ Gehring logrou en 1994 a mutación da sobreexpresión do xene Pax6, creando estruturas oculares ectópicas que expresarían o xene Pax6. Dous dos seus colaboradores, George Hager e Patrick Callaerts, utilizaron o factor de transcrición Gal14 de fermentos paira transferir o eyeless cDNA a estruturas fose do disco de ollo.
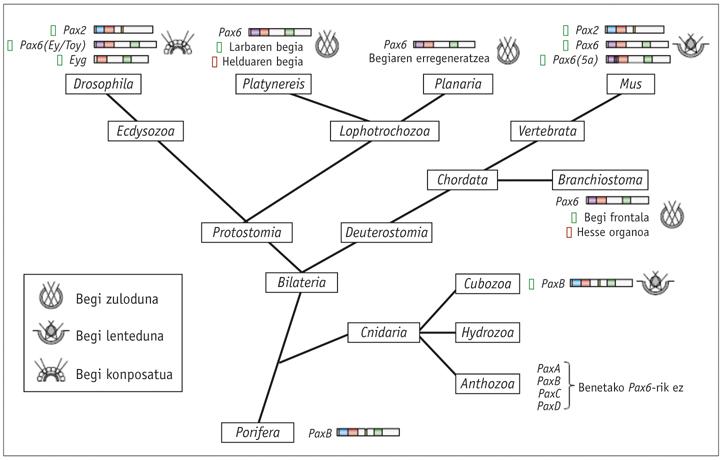
Os dous colaboradores lograron crear estruturas visuais ectópicas en antenas, patas e ás. Posteriormente, mediante retinogramas demostrouse que algúns dos ollos que aparecían nas antenas eran totalmente funcionais (Halder et ao. , 1995). Así mesmo, observaron que o xene Pax6 do rato era capaz de inserilo no Drosophila para producir ollos ectópicos (Gehring et ao. , 1994).
Estes experimentos demostraron que Pax6 era o principal xene de control e que este xene "estimulante" podía dar inicio á morfogénesis do ollo, tanto en mamíferos como en insectos.
Os xenes homólogos de Pax6 atopáronse en todos os animais bilaterais estudados, desde o planeta ao home, incluíndo C. elegans. Realizáronse probas con diferentes animais e comprobouse que o xene Pax6 de todos eles é capaz de inducir os ollos ectópicos en Drosophila. A única excepción desta lei é a tigrina planaria Dugesia, que se afastou moito. En canto aos cnidarios, só algúns hidrozoos teñen os ollos e non se sabe si o resto das medusas perderon os seus ollos ao longo da evolución ou nunca os tiveron.
Visión de cores
Hai estudos que demostran que o sistema de visión en cor é una cuestión ancestral, xa que desde o principio existían un pigmento visual (chamado S) moldeado paira absorber a luz a 500 <nm e outro (L) adaptado a 500 > nm (Mollon, 1989). A rodopsina, un pigmento tan antigo como estes, ten una capacidade de absorción máxima duns 500 nm e non ten ningún efecto na visión en cor. En xeral, os pigmentos necesarios paira a visión da cor atópanse nos conos fotorreceptores, sendo funcionais unicamente baixo a influencia da luz. Doutra banda, a rodopsina atópase nos paus fotorreceptores e traballa na escuridade.
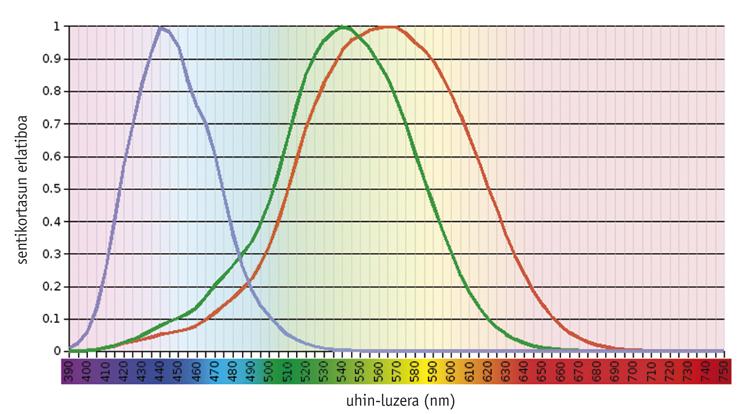
Os vertebrados actuais mostran una gran variabilidade na súa capacidade visual, desde a densidade e distribución espacial dos diferentes tipos de conos até a absorción máxima dos pigmentos dos conos (Yokoyama, 1998). Nun dos extremos, a maioría dos mamíferos só conteñen tres pigmentos: dous pigmentos precursores de cono e rodopsina. No outro extremo evolutivo, os pitos teñen seis pigmentos.
Segundo estudos realizados nos vertebrados máis antigos, como os fuxidos, o precursor común dos teleosteos e os amniotas tiña catro tipos de fotopigmentos, do mesmo xeito que as aves e réptiles actuais (Bowmaker, 1998). Con todo, crese que debido ao carácter nocturno dos mamíferos prematuros perdéronse dous fotopigmentos ao longo da evolución, e a maioría dos mamíferos euterio actuais só manteñen fotopigmentos S e L, é dicir, son dicromáticos.
Os seres humanos e os seus primates máis próximos mostran un modelo de complexidade media. Os seres humanos teñen catro pigmentos visuais: Un só membro da familia S (azul), dúas fotopigmentos da familia L (verde ou LM e vermello ou LL) e rodopsina.
A visión tricromática é de gran importancia paira os primates e relacionouse coa vantaxe de atopar comida nos bosques. As obras máis recentes demostraron a importancia ecolóxica da trikromacia comparando a súa capacidade de adaptación ao bosque de monos bicromáticos e tricromáticos. En todos os primates, o fotopigmento S é codificado por un xene autosómico, mentres que o fotopigmento L está codificado no cromosoma X. Nos primates antropoidea e monos ularios do Vello Mundo, a tricromacia xorde pola duplicación do xene L, dando lugar ao xene LM. Con todo, na maioría dos primates do Novo Mundo, o cromosoma X é o único xene da familia L. Na medida en que o xene está asociado a X, as femias heterocigóticas son tricromáticas, e as femias homocigóticas e machas, dicromáticas (cegas de verde/vermello) (Jacobs, 1998).
BIBLIOGRAFÍA
Zu idazle
Zientzia aldizkaria

