

Begiari begira
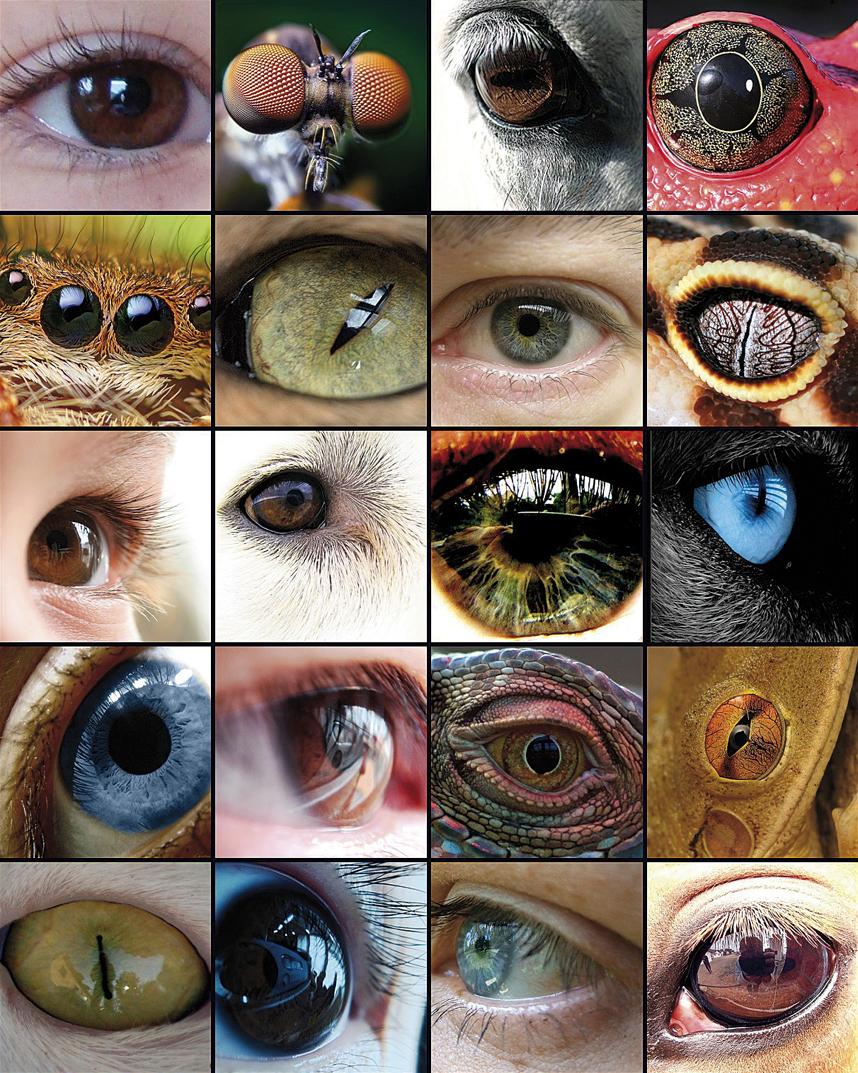
Begiaren eboluzioa ikerketa ugariren oinarri izan da, taxoi ezberdinetan dagoen organo homologoaren eredu adierazgarria baita. Begiaren zenbait osagaik, esaterako, ikusmen-pigmentuek, aitzindari komuna dutela ematen duen arren, irudi konplexuak sortzeko gai diren begiak 50-100 aldiz sortu dira eboluzioan, behin eta berriz proteina eta gene berdinez baliatuz.
Charles Darwinek berak Espezieen jatorria (1882) liburuan aitortzen zuen zailtasun handiak zituela begiaren eboluzioa azaltzeko, eta kapitulu oso bat eskaini zion begiari Teoriaren zailtasunak izenburupean, zera esanez: "begia moduko organo konplexu bat hautespenez sortu izanak, benetan diot, izugarrizko erokeria dirudi". Baina gero honela jarraitzen zuen: "Horregatik, baldin begi sinple eta inperfektu batetik gaur egungo begi perfektu eta konplexua sortzeko adina aldaketa gertatu bazen, benetan gertatu izan diren bezala, eta aldaketa horien eramaileak ziren animaliei erabilgarriak suertatu bazitzaizkien bizitzaren ingurune aldakorraren aurrean, orduan gure begi konplexu perfektuak hautespen naturalez sortu izanak, gure irudimena gainditzen badu ere, ez dirudi horren ideia iraultzailea".
Zelula fotorrezeptoreen jatorria metazooetan
Zelula fotorrezeptoreak fototransdukzioa gauzatzeko gai diren erretinako nerbio-zelulak dira. Fotorrezeptoreak garrantzi biologiko handikoak dira, fotoiak xurgatuz, eta bidezidor biokimiko konplexu ugari eginda, jasotako seinale hori irudi bilaka daitekeelako burmuinean.
Bi hipotesi daude metazooetako zelula fotorrezeptoreen jatorria azaltzeko: lehenengoa desberdintzapenean oinarritzen da, eta bigarrena, sinbiosian. Lehen hipotesiaren arabera, metazooak flagelatu-kolonia batetik abiatuta sortu ziren, eta zelula guztiek hasieratik organulu fotorrezeptore bat zuten. Organulu fotorrezeptore horrek begi-orbain modura jarduten zuen, eta seinaleak flageloan zehar transmititzen zituen fototaxiaren aurreko erantzun gisa.

Sinbiosiaren hipotesiaren arabera, berriz, Volvox edo Chlamydomonas moduko flagelatuetan dute jatorria; haietan, organulu fotosentikorra kloroplastoetan dago, eta horrek iradokitzen du argiaren pertzepzioa zianobakterioetatik aurrerako eboluzioan dagoela, eta gero kloroplasto modura integratu zela zelula eukariotoetan ere. Greuet-ek azaldu zuen 1965ean hipotesi horren aldeko beste froga bat, Erythropsis eta Warnovia moduko dinoflagelatuen organulu fotorrezeptoreak deskribatzean, zeinak giza begia bezain garaturik baina zelula bakarraren barnean agregaturik baitaude; kornearen antzeko gainazal bat dute, lenteen antzeko egitura bat, erretinaren antzeko beste bat eta pigmentu-organuluak. Dinoflagelatuak koralen, itsas anemonen eta beste knidario batzuen sinbionte arruntak direla-eta, agian argiari erantzuteko beharrezko geneak transferitu zizkieten knidarioei; horrek esplikatuko luke knidarioei begia bat-batean agertu izana.
Sinbionteen hipotesiaren arabera, argiarekiko sentikortasuna zianobakterioetan sortu zen lehendabizi. Zianobakterio horiek alga gorrien zelula eukariotikoek barneratu zituzten, kloroplasto primario modura. Gero, alga gorriak dinoflagelatuekin sinbiosian egotera pasatu ziren, bigarren mailako kloroplasto gisa. Zenbait dinoflagelatu-espezietan, Erythropsis eta Warnovia kasurako, ez dira aurkitu bigarren mailako kloroplastoak, eta uste da horiek eboluzionatu egin zutela eta organulu fotorrezeptore eraginkor bihurtu, Greuet-ek iradoki bezala. Azkenik, eta dinoflagelatuak knidarioen sinbionte arruntak izanik, fotorrezeptoreen geneak helarazi zizkieten haiei. Azken hori da ereduaren urrats zalantzazkoena, nahiz eta dinoflagelatuen hainbat gene aurkitu den knidarioen genoman txertaturik.
Begiaren garapenaren analisia
Animalietan, erraz detekta daitezke begiaren garapenean eragina duten mutazioak, eta Drosophila eulian gertatzen den eyeless (ey) mutazioa 1915ean aurkitu zuen lehen aldiz Hoge ikertzaileak. Saguetan antzeko mutazio bat aurkitu zen, eta small eye izendatu, animalia heterozigotoek begiak oso murriztuta zituztelako; uteroan hiltzen ziren homozigotoen fetuek, aldiz, begiak falta zituzten, baina baita sudurra eta burmuineko zenbait zati ere (Hill et al ., 1991).
Gizakian, oso antzeko fenotipoa sortzen du aniridia izeneko sindrome hereditarioak. Small eye eta aniridia geneak Walther-ek eta Gruss-ek (1991), eta Ton eta haren taldeak (1991) klonatu zituzten, hurrenez hurren, eta ikusi zuten biek izugarri kontserbatuta zegoen Pax6 geneari erantzuten ziotela. Quiring eta haren taldeak (1994) klonatu zuten Drosophilaren Pax6 homologoa, eta harrituta ikusi zuten Hoge-ren eyeless (ey) genearen berdina zela. Small eye, aniridia eta eyeless gene homologoen mutazioak izateak iradokitzen zuen Pax6 genea zela begiaren garapenaren kontrol-gene nagusia ornodun zein ornogabeetan.
Hipotesi hori frogatzeko, WJ Gehring ikertzaileak Pax6 genearen gainespresioaren mutazioa lortu zuen 1994an, Pax6 genea espresatuko zuten begi-egitura ektopikoak sortuz. Haren kolaboratzaileetako bik, George Halder-ek eta Patrick Callaerts-ek, legamien gal14 transkripzio-faktorea erabili zuten eyeless cDNA begi-diskotik kanpoko egituretara bideratzeko.
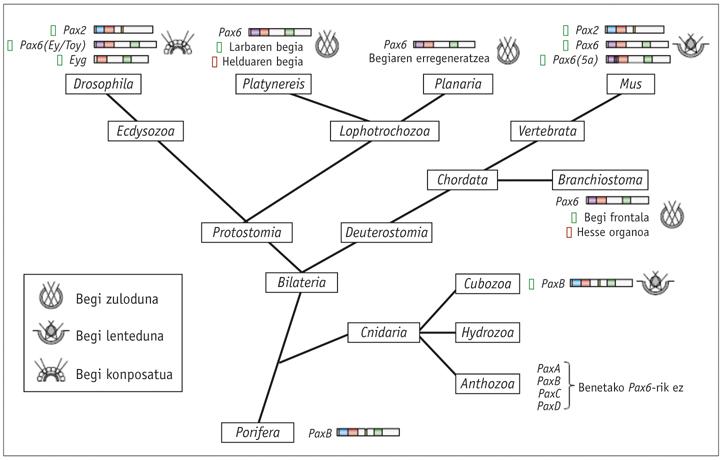
Bi kolaboratzaileok lortu zuten begi-egitura ektopikoak antena, hanka eta hegoetan sortzea. Geroago, erretinogramen bidez frogatu zen antenetan agertzen ziren begietako batzuk guztiz funtzionalak zirela (Halder et al. , 1995). Era berean, saguaren Pax6 genea Drosophilan txertatu eta han begi ektopikoak sortzeko gai zela ere behatu zuten (Gehring et al. , 1994).
Esperimentu horiek frogatu zuten Pax6 kontrol-gene nagusia zela, eta gene "pizgarri" horrek begiaren morfogenesiari hasiera eman ziezaiokeela bai ugaztunetan bai intsektuetan.
Pax6 -ren gene homologoak aztertutako animalia bilateral guztietan aurkitu izan dira, planarietatik gizakietara, C. elegans barne. Hainbat animaliarekin frogak egin dira, eta ikusi da guztien Pax6 genea begi ektopikoak induzitzeko gai dela Drosophilan . Lege horretako salbuespen bakarra Dugesia tigrina planaria da, antzaz izugarri aldendu dena. Knidarioei dagokienez, soilik zenbait hidrozook dituzte begiak, eta ez da ezagutzen ea gainerako marmokek beren begiak galdu dituzten eboluzioan zehar edo ez dituzten inoiz izan.
Koloretako ikusmena
Zenbait ikerketaren arabera, koloretako ikusmen-sistema antzinako kontua da, hasieratik baitzeuden <500 nm-ra argia xurgatzeko moldaturiko ikusmen-pigmentu bat (S deiturikoa) eta >500 nm-ra moldatutako beste bat (L) (Mollon, 1989). Errodopsinak, horiek bezain zaharra izango litzatekeen pigmentuak, xurgapen-ahalmen maximoa 500 nm inguruan du, eta ez du inolako eraginik koloretan ikusteko ahalmenean. Oro har, kolorea ikusteko beharrezko diren pigmentuak kono fotorrezeptoreetan daude, eta argiaren eraginpean soilik dira funtzionalak. Bestalde, errodopsina makil fotorrezeptoreetan dago, eta iluntasunean dihardu.
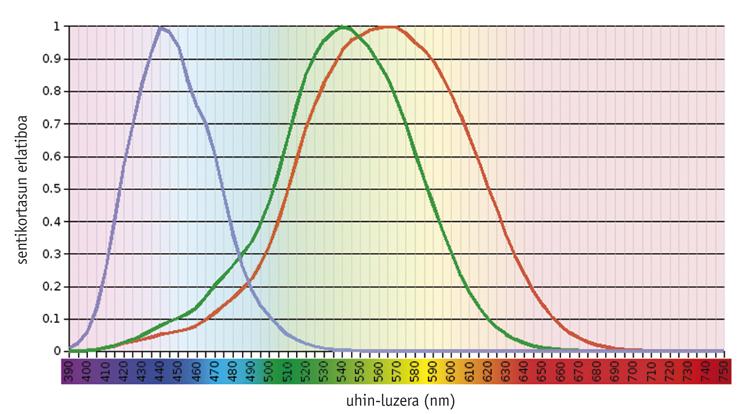
Gaur egungo ornodunek aldakortasun handia erakusten dute beren ikusmen-ahalmenean, hasi kono-moten dentsitate eta banaketa espazialetik, konoen pigmentuetako absortzio maximoraino (Yokoyama, 1998). Mutur batean, ugaztun gehienek hiru pigmentu baino ez dituzte: bi kono-pigmentu aitzindariak eta errodopsina. Beste mutur ebolutiboan, oilaskoek sei pigmentu dituzte.
Ornodun zaharrenetan egindako ikerketen arabera (gaizkatetan, esate baterako), teleosteoen eta amniotoen aitzindari komunak lau fotopigmentu-mota zituen, gaur egungo hegazti eta narrastiek bezala (Bowmaker, 1998). Hala ere, uste da ugaztun goiztiarren izaera gautarra zela-eta eboluzioan zehar bi fotopigmentu galdu zirela, eta gaur egungo ugaztun euterio gehienek soilik S eta L fotopigmentuak mantentzen dituzte, hau da, dikromatikoak dira.
Gizakiek eta haien primate hurbilenekoenek konplexutasun ertaineko eredu bat erakusten dute. Gizakiek lau ikusmen-pigmentu dituzte: S familiako kide bakarra (urdina), L familiako bi fotopigmentu (berdea edo LM deitutakoa, eta gorria edo LL) eta errodopsina.
Ikusmen trikromatikoa garrantzi handikoa da primateentzat, eta basoetan jana bilatzeko abantailarekin lotu izan da. Lan berrienek trikromaziaren garrantzi ekologikoa frogatu dute, tximino bikromatikoen eta trikromatikoen basoan moldatzeko gaitasuna alderatuz. Primate guztietan, S fotopigmentua gene autosomiko batek kodetzen du; L fotopigmentua, aldiz, X kromosoman kodeturik dago. Mundu Zaharreko primate antropoideoetan eta tximino ulularietan, trikromazia L genearen duplikazioz sortu da, LM genea sortuz. Hala ere, Mundu Berriko primate gehienetan X kromosoman L familiako gene bakarra dago. Genea X-ri loturik dagoen heinean, eme heterozigotoak trikromatikoak dira, eta eme homozigotoak eta arrak, dikromatikoak (berde/gorrirako itsuak) (Jacobs, 1998).
BIBLIOGRAFIA
Idatzi zuk zeuk Gai librean atalean
Gai librean aritzeko, bidali zure artikulua aldizkaria@elhuyar.eus helbidera
Hauek dira Gai librean atalean Idazteko arauak
Zu idazle
Zientzia aldizkaria


