

Mirant a l'ull
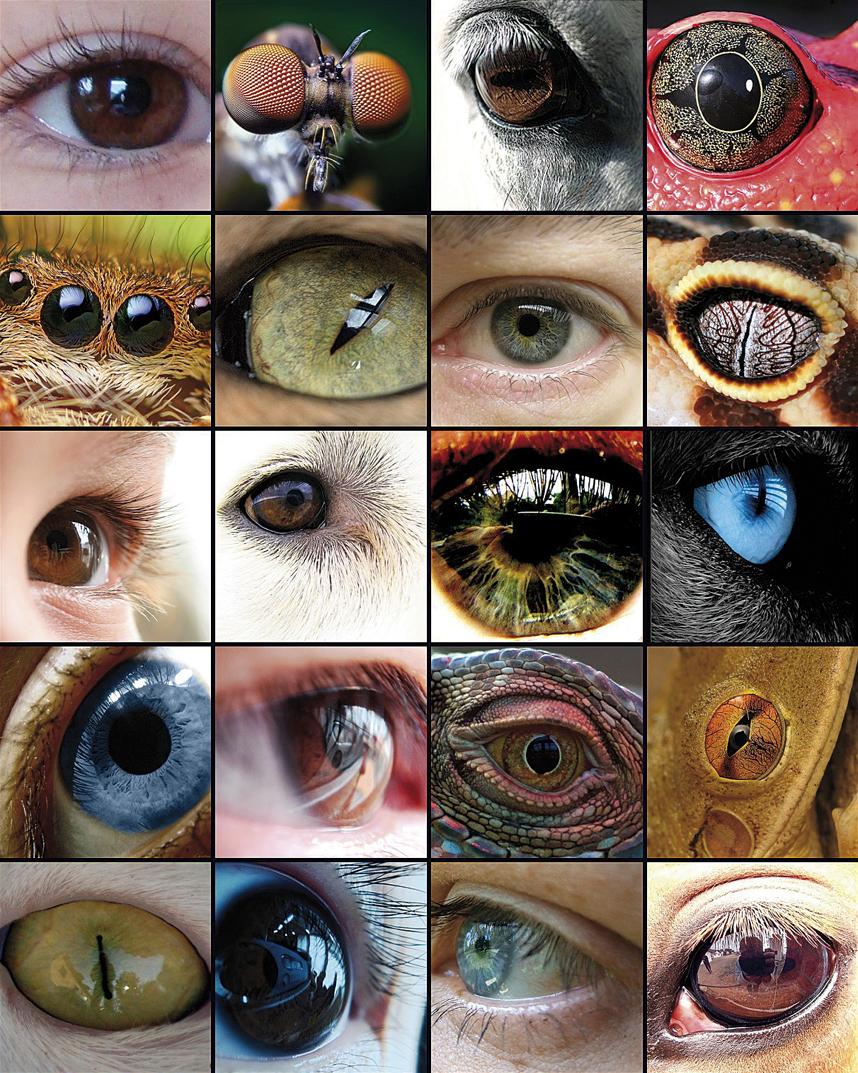
L'evolució de l'ull ha estat la base de nombrosos estudis, ja que és un exemple representatiu de l'òrgan homòleg en els diferents tàxons. Encara que alguns components de l'ull, com els pigments visuals, semblen tenir un precursor comú, els ulls capaços de produir imatges complexes han evolucionat entre 50 i 100 vegades, recorrent a les mateixes proteïnes i gens de manera recurrent.
El mateix Charles Darwin reconeixia en el seu llibre L'origen de les espècies (1882) que tenia grans dificultats per a explicar l'evolució de l'ull i va dedicar un capítol complet a l'ull sota el títol Les dificultats de la teoria dient: "el fet que un òrgan tan complex com l'ull hagi estat creat per elecció sembla, certament, una bogeria enorme". Però després seguia així: "Per això, si d'un ull simple i imperfecte es van produir els canvis necessaris per a crear l'ull perfecte i complex d'avui, tal com s'han produït en realitat, i si als animals portadors d'aquests canvis els resultaven útils davant l'entorn canviant de la vida, el fet que els nostres ulls complexos perfectes es formessin per selecció natural, encara que superi la nostra imaginació, no sembla una idea tan revolucionària".
Origen de les cèl·lules fotorreceptoras en els metazoos
Les cèl·lules fotorreceptoras són cèl·lules nervioses de la retina capaces de dur a terme la fototransducción. Els fotorreceptores són de gran importància biològica mitjançant l'absorció de fotons i la realització de nombroses senderes bioquímiques complexos que converteixen el senyal rebut en imatge en el cervell.
Existeixen dues hipòtesis per a explicar l'origen de les cèl·lules fotorreceptoras dels metazoos: la primera es basa en la diferenciació i la segona en la simbiosi. Segons la primera hipòtesi, els metazoos es van formar a partir d'una colònia de flagel·lats, i totes les cèl·lules tenien des del principi un orgáculo fotoreceptor. Aquest orgànuls fotoreceptor actuava com a cicatriu ocular i com a resposta a la fototaxis que transmetia els senyals a través del flagel.

Segons la hipòtesi de la simbiosi, tenen el seu origen en flagel·lats com Volvox o Chlamydomonas, en els quals l'orgáculo fotosensible es troba en cloroplastos, la qual cosa suggereix que la percepció de la llum es troba en una evolució a partir de les cianobacterias i que posteriorment s'integra com a cloroplast en cèl·lules eucariotes. Una altra prova a favor d'aquesta hipòtesi, va explicar Greuet en 1965, en descriure els orgànuls fotorreceptores de dinoflagelados com Erythropsis i Warnovia, tan desenvolupats com l'ull humà però agregats a l'interior d'una sola cèl·lula, amb una superfície similar a la còrnia, una estructura similar a les lents, una altra similar a la retina i orgànuls pigments. Els dinoflagelados, a causa dels simbionts comuns de corals, anemones marines i uns altres cnidarios, podien haver transferit als cnidarios els gens necessaris per a respondre a la llum, la qual cosa explicaria l'aparició sobtada de l'ull als cnidarios.
Segons la hipòtesi dels Sinbiontes, la sensibilitat a la llum va sorgir primer en les cianobacterias. Aquestes cianobacterias van ser interioritzades per cèl·lules eucariotes d'algues vermelles com a cloroplastos primaris. Posteriorment, les algues vermelles van passar a estar en simbiosis amb dinoflagelados com a cloroplastos secundaris. En algunes espècies de dinoflagelados, com Erythropsis i Warnovia, no s'han trobat cloroplastos secundaris que es creu que van evolucionar i es van convertir en orgànuls fotorreceptores efectius, com suggereix Greuet. Finalment, i atès que els dinoflagelados són sinbiontes comuns als cnidarios, van transmetre a aquests els gens dels fotorreceptores. Aquest últim és el pas més incert del model, encara que s'han trobat diversos gens dels dinoflagelados integrats en el genoma dels cnidarios.
Anàlisi del desenvolupament de l'ull
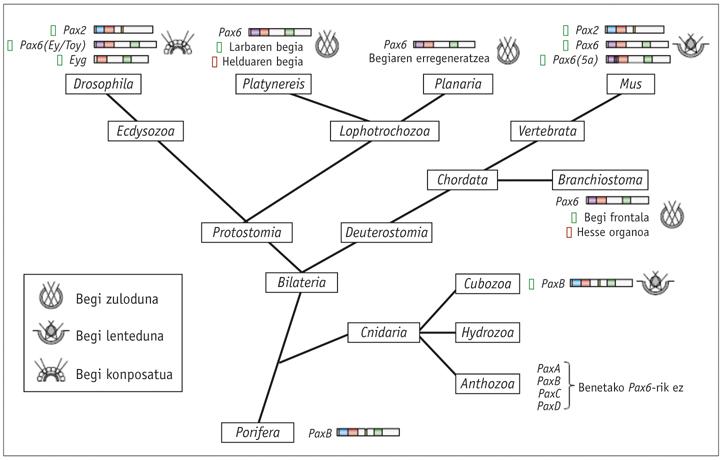
En els animals és fàcil detectar mutacions que afecten el desenvolupament de l'ull i la mutació eyeless (ey) en la mosca Drosophila va ser descoberta per primera vegada en 1915 per l'investigador Hoge. En el ratolí es va trobar una mutació semblant i es va denominar small eye perquè els animals heterocigóticos tenien els ulls molt reduïts, mentre que els fetus d'homocigótanos que morien en l'úter faltaven els ulls, però també el nas i algunes parts del cervell (Hill et al ., 1991).
En l'home, una síndrome hereditària anomenada aniridia produeix un fenotip molt semblant. Els gens Small eye i aniridia van ser clonats per Walther i Gruss (1991) i Ton i el seu grup (1991) respectivament, i van veure que tots dos responien al gen Pax6 extremadament conservat. Quiring i el seu equip (1994) van clonar a l'homòleg Pax6 de Drosophila i van veure sorprès que l'eyeless (ey) d'Hoge era igual al gen. L'existència de mutacions de gens homòlegs Small eye, aniridia e eyeless indicava que el gen Pax6 era el principal gen de control del desenvolupament ocular tant en vertebrats com invertebrats.
Per a demostrar aquesta hipòtesi, l'investigador WJ Gehring va aconseguir en 1994 la mutació de la sobreexpresión del gen Pax6, creant estructures oculars ectòpiques que expressarien el gen Pax6. Dos dels seus col·laboradors, George Hager i Patrick Callaerts, van utilitzar el factor de transcripció Gal14 de llevats per a transferir l'eyeless cDNA a estructures fora del disc d'ull.
Els dos col·laboradors van aconseguir crear estructures visuals ectòpiques en antenes, potes i ales. Posteriorment, mitjançant retinogramas es va demostrar que alguns dels ulls que apareixien en les antenes eren totalment funcionals (Halder et al. , 1995). Així mateix, van observar que el gen Pax6 del ratolí era capaç d'inserir-lo en el Drosophila per a produir ulls ectòpics (Gehring et al. , 1994).
Aquests experiments van demostrar que Pax6 era el principal gen de control i que aquest gen "estimulant" podia donar inici a la morfogènesi de l'ull, tant en mamífers com en insectes.
Els gens homòlegs de Pax6 s'han trobat en tots els animals bilaterals estudiats, des del planeta a l'home, incloent C. elegans. S'han realitzat proves amb diferents animals i s'ha comprovat que el gen Pax6 de tots ells és capaç d'induir els ulls ectòpics en Drosophila. L'única excepció d'aquesta llei és la tigrina planaria Dugesia, que s'ha allunyat molt. Quant als cnidarios, només alguns hidrozoos tenen els ulls i no se sap si la resta de les meduses han perdut els seus ulls al llarg de l'evolució o mai els han tingut.
Visió de colors
Hi ha estudis que demostren que el sistema de visió en color és una qüestió ancestral, ja que des del principi existien un pigment visual (anomenat S) modelat per a absorbir la llum a 500 <nm i un altre (L) adaptat a 500 > nm (Mollon, 1989). La rodopsina, un pigment tan antic com aquests, té una capacitat d'absorció màxima d'uns 500 nm i no té cap efecte en la visió en color. En general, els pigments necessaris per a la visió del color es troben en els cons fotorreceptores, sent funcionals únicament sota la influència de la llum. D'altra banda, la rodopsina es troba en els pals fotorreceptores i treballa en la foscor.
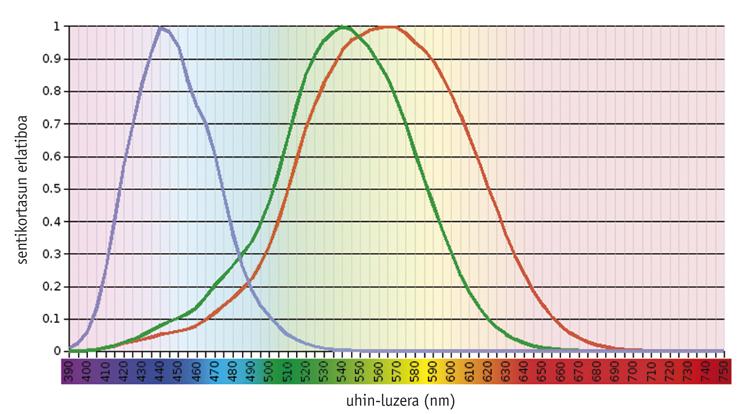
Els vertebrats actuals mostren una gran variabilitat en la seva capacitat visual, des de la densitat i distribució espacial dels diferents tipus de cons fins a l'absorció màxima dels pigments dels cons (Yokoyama, 1998). En un dels extrems, la majoria dels mamífers només contenen tres pigments: dos pigments precursors de con i rodopsina. En l'altre extrem evolutiu, els pollastres tenen sis pigments.
Segons estudis realitzats en els vertebrats més antics, com els facinerosos, el precursor comú dels teleosteos i els amniotas tenia quatre tipus de fotopigmentos, igual que els ocells i rèptils actuals (Bowmaker, 1998). No obstant això, es creu que a causa del caràcter nocturn dels mamífers prematurs es van perdre dos fotopigmentos al llarg de l'evolució, i la majoria dels mamífers euterio actuals només mantenen fotopigmentos S i L, és a dir, són dicromáticos.
Els éssers humans i els seus primats més pròxims mostren un model de complexitat mitjana. Els éssers humans tenen quatre pigments visuals: Un sol membre de la família S (blau), dos fotopigmentos de la família L (verd o LM i vermell o LL) i rodopsina.
La visió tricromàtica és de gran importància per als primats i s'ha relacionat amb l'avantatge de trobar menjar en els boscos. Les obres més recents han demostrat la importància ecològica de la trikromacia comparant la seva capacitat d'adaptació al bosc de micos bicromáticos i tricromàtics. En tots els primats, el fotopigmento S és codificat per un gen autosómico, mentre que el fotopigmento L està codificat en el cromosoma X. En els primats antropoidea i bufons ularios del Vell Món, la tricromacia sorgeix per la duplicació del gen L, donant lloc al gen LM. No obstant això, en la majoria dels primats del Nou Món, el cromosoma X és l'únic gen de la família L. En la mesura en què el gen està associat a X, les femelles heterocigóticas són tricromàtiques, i les femelles homocigóticas i gallimarsots, dicromáticas (cegues de verd/vermell) (Jacobs, 1998).
BIBLIOGRAFIA
Zu idazle
Zientzia aldizkaria

