

Si Deus non foi… Entón, cal foi?

No número de setembro de 2012 publicouse un artigo "mugalari" na revista Elhuyar. Digo "limitante" porque o tema do artigo atravesaba as fronteiras de certos ámbitos de coñecemento: relixión, anatomía humana, tradución, mitoloxía... O tema do artigo foi analizar como una carencia do home, una característica anatómica moi estendida entre os mamíferos, a falta de óso do pene, podería explicarse. Na linguaxe técnica da anatomía, o óso do pene denomínase latín vos penis ou baculum (o pau deriva da mesma palabra latina).
Na explicación mitológica do porqué desta carencia requiríase a intervención de Deus. Pareceume interesante analizar o tema desde o punto de vista científico. Pero neste caso, cando Napoleón preguntou a Laplace como o fixo no gran libro Traité de Mécanique Céleste paira explicar o funcionamento do universo sen mencionar nunca ao Creador, vou traer a resposta de Laplace a Napoleón: Je n'avais pas besoin de cette hypothèse-là ("Non necesitei esta hipótese"). É dicir, eu tampouco necesitarei a Deus paira explicar cientificamente a falta de óso do pene. Iso si, necesitarei atravesar certas limitacións, como saír do campo da bioloxía molecular e entrar no amplo campo da teoría da evolución. Por tanto, este artigo tamén será necesariamente "limitativo".
Na pregunta do título deste artigo ("cales..?") como implicitamente menciónase a unha persoa, despois de mencionar que non terei necesidade de Deus paira a explicación, creo que chegou o momento de reescribir a pregunta, porque cientificamente será máis correcto preguntar: Entón, que provocou a perda do óso do pene? Polo que sabemos en xaneiro de 2013, probablemente porque con certeza absoluta na ciencia apenas se pode saber nada, a resposta é tan exacta como seca: os delitos dunha das secuencias de intensificadores que se atopan xunto ao xene do receptor de andrógenos e que se conservaron moi ben durante a evolución. No ámbito da xenética, a perda de material xenético coñécese como delezio. Recentemente (marzo de 2011) publicouse que desde que a tribo Hominini separouse da tribo Panini (chimpancés), producíronse 510 delitos ou perdas de secuencia ao longo da ruta evolutiva que nos levou ao ser humano. Una das partes do cromosoma perdidas levaba a orde de formar o óso do pene nun momento dado do desenvolvemento embrionario. Por que non temos espiñas de pene? A resposta sintética é que nalgún momento produciuse un delito. Una vez aclarado cal pode ser a razón concreta, parece que o tema estaría esgotado, pero como estar a preguntar constantemente polo porqué é a forma na que a ciencia avanza, o núcleo deste artigo será tratar de responder á seguinte pregunta profunda e grosa.
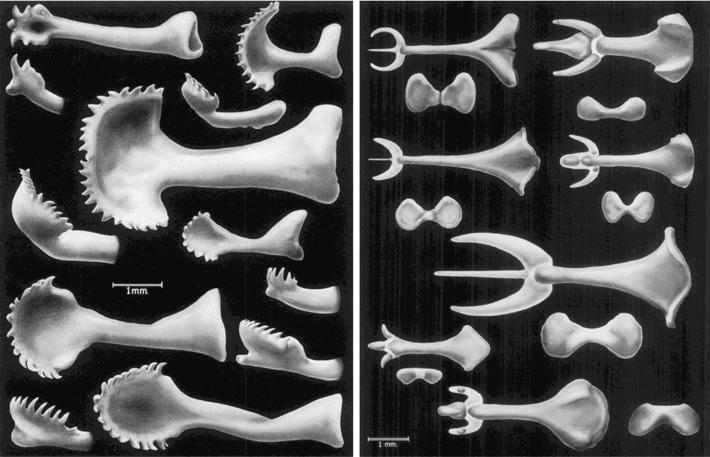
Como se pode explicar, pois, que ao longo da evolución sexa un home sen espiñas? Que vantaxe habería de ter? Na última figura pódese observar que morfológicamente o óso do pene é moi plástico, cunha gran riqueza de aspecto. Esa riqueza indicaríanos, desde o punto de vista reprodutivo, que o óso do pene cumpre una función moi importante, que nesta pequena parte da súa anatomía o macho xoga moito... Ante leste polimorfismo, parece máis urxente, por tanto, dar una explicación adecuada e convincente á elección dun home sen espiñas de pene.
A elección dun home sen espiñas de pene non foi una labor de selección natural, xa que esta característica anatómica non achega en principio ningunha vantaxe na loita pola vida. O avance deste tipo de homes ao longo da evolución débese á selección sexual, é dicir, aínda que pareza sorprendente, o pene deshuesado aumentou o éxito reprodutivo do home, axudándolle a triunfar na loita pola reprodución. Non é evidente, paira a maioría, como e por que.
A selección sexual é una competición intraespecífica --é dicir, una competición entre individuos da mesma especie (no noso caso animais) paira ter opción de reprodución, na que dunha ou outra maneira participan ambos os sexos. A selección pode ser intrasexual, é dicir, que a selección se produza como consecuencia da competencia entre unidades do mesmo sexo, aínda que normalmente. A competición pode consistir nunha cúpula previa (pelexas entre machos adultos) ou postcúpula (competición entre esperas). Doutra banda, a selección pode ser tamén intersexual, é dicir, un sexo pode elixir entre os números do outro sexo. Ninguén se sorprenderá si le que o sexo máis selector é o máis prexudicado en caso de mala elección. Normalmente as femias son selectoras estritas e os machos, seleccionados. Tamén neste caso, a selección pode producirse antes da copulación, por exemplo na elección de parella, coñecida como selección clásica da femia. E pode suceder despois de copular. Nos animais que se reproducen por fecundación interna, a selección intersexual que se produce dentro do corpo da femia denomínase selección críptica da femia, xa que o macho non sabe como actúa (elixe ou rexeita) cos seus espermas.
Analicemos en cal destes catro casos da selección sexual fose una vantaxe a perda do óso do pene, esperando atopar así o motivo da elección dun home sen espiñas. Nas competicións intrasexuales, é dicir, nas competicións con outros homes, non parece que a perda do óso do pene ofreza ningunha vantaxe, nin nas competicións prenatales nin nas posteriores. Tamén desde o punto de vista da selección clásica da femia, non se aprecia ningunha vantaxe na perda do óso do pene; neste último caso, parece que á contraria pódeselle atopar una explicación máis sinxela. Por tanto, só no campo da selección críptica da femia poderemos buscar e atopar a explicación.
Nos animais que se reproducen sexualmente, é imposible evitar a aparición de conflitos entre os intereses de ambos os sexos, xa que os investimentos enerxéticos de ambos os sexos son moi diferentes nos asuntos reprodutivos. Como consecuencia destes conflitos, moitas das características biolóxicas han cambiado na evolución da nosa especie. Con todo, a colaboración entre ambas as parellas, polo menos en certa medida, é imprescindible. En calquera caso, a colaboración entre ambos os sexos materializarase no avance do produto reprodutivo, é dicir, no coidado e reprodución das crías.
Una das características máis características da nosa especie é o tamaño do noso cerebro, cun peso aproximado de 1,5 kg, entre 3 e 4 veces maior que o do noso parente filogenético máis próximo, o do chimpancé. A aparición do xénero Homo indícanos que fai uns 2 millóns de anos produciuse algo que supuxo un aumento do tamaño do noso cerebro. Podería deberse a un dos 510 delitos mencionados ao comezo deste traballo, xa que una delincuencia pode ocasionar a perda dunha estrutura (o óso do pene) e a ampliación dunha estrutura por outra. A perda do GADD45G está relacionada co aumento de determinados lugares específicos do cerebro humano, aínda que aínda non sabemos si existe relación causa-efecto entre ambos os factores.
Ao nacer, o tamaño do cerebro dun ser humano só representa o 30% do seu tamaño cando alcanza a súa madurez. O cerebro do chimpancé recentemente nado é o 40% do tamaño do cerebro do gran adulto ao nacer. En canto ao desenvolvemento do cerebro, dado que os nosos fillos nacen máis "inmaturos" que os dos chimpancés, as necesidades de coidado postparto pasaron a ser moito maiores. A partir do momento en que o cerebro comezou a crecer de forma acelerada na evolución humana, os intereses reprodutivos da muller cambiaron considerablemente. A partir de entón, o obxectivo da muller non era outro que copular cun alfa macho impregnado de testosterona na mesquita dos bos xenes, senón que o seu principal froito reprodutivo sería atopar a un home máis feminizado, que tamén realizase as tarefas que lle corresponderían a unha femia, e que, no que respecta ao crecemento do neno, ademais de contribuír, asumise esta tarefa. Paira lograr este obxectivo estratéxico foi necesario que se producisen algúns cambios anatómicos, fisiológicos e etológicos na muller.
Ademais do gran tamaño do cerebro, entre os primates hai una característica totalmente inusual e propia: a ovulación críptica da muller, é dicir, que a muller non reflicta ningún sinal visible da data en que é fértil. Con esta estratexia reprodutiva da muller, o home que coidaba e alimentaba á súa parella de nais -- nenos nunca podía estar seguro de que o neno que estaba a coidar era realmente seu, xa que non sabía se a muller coa que se reproducía puido ou non ter sexo. Máis aínda, paira aumentar a variabilidade xenética e reducir ao mínimo os efectos perniciosos da consanguinidad, como ocorre nos nosos parentes máis próximos, os bonobos, cando a estratexia recompensada evolutivamente nos animais que habitan en pequenos grupos é a poliginandría --poligamia reducida-.
A saúde é una característica de ser bos xenes, que significa que temos una gran capacidade de avanzar nas condicións nas que vivimos. Así, coa esperanza de ter fillos sans, en primeiro lugar, a muller buscaba parella sa paira manter relacións sexuais. Como o pene sen ósos baséase nun sistema hidráulico de bombeo, o fallo na tentación do pene é una sinal ou aviso moi sensible, honesto e precoz que indica á muller que o home padece algunha enfermidade física ou mental. No perfil de Facebook podemos mentir. Niso non. Non hai que dicilo, coa intención directa de atopar un home san que vivise o suficiente como paira coidar do seu fillo, indirectamente e de maneira inconsciente, a muller realizou una selección sexual que permitiu aos vendedores de Viagra facer negocio.
Nesta viaxe evolutiva até o pene sen ósos do home, seguro que a muller, a nosa deusa, foi o vento que mantivo as velas constantemente inchadas.
Bibliografía
Zu idazle
Zientzia aldizkaria

- Babesleak
-

-

Elhuyar
Nor gara | Kontaktua |
Publizitatea
| Laguntza
Pribatutasun politika | Cookien politika
ISSN-2603-6614 Elhuyar