

Grans Misteris de les Cèl·lules Petites I com els Indicadors Fluorescents s'han convertit en informació
Els descobriments d'última generació han donat la volta i mitja al món. D'una banda, les recerques orientades a l'espai ens han portat a l'exterior de la terra i a altres planetes. La idea que en l'Univers han existit o existeixen altres formes de vida és avui més forta que mai. D'altra banda, ha aprofundit en els treballs sobre l'estructura de les partícules internes de l'àtom de viatge a l'interior, que també han aportat noves intencions sobre l'univers. No obstant això, encara són més les preguntes pendents que les respostes.
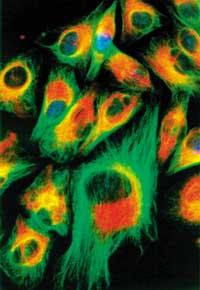
La cèl·lula viva és sens dubte l'estructura més complexa i sorprenent del nostre planeta. D'una banda, per la seva rica informació interna i la seva estructura compacta (pensem, per exemple, que el programa de desenvolupament per a la creació d'una balena s'emmagatzema en pocs nanogramos d'ADN). D'altra banda, cada cèl·lula exerceix un treball complex en un lloc molt petit. En tot moment, milers de molècules diferents estan duent a terme nombroses reaccions bioquímiques dins d'un pla metabòlic concret i catramílico. Al mateix temps, mantenen l'estructura cel·lular i, a més, compleixen correctament la seva funció en un univers de cèl·lules de tot l'organisme (el nombre de cèl·lules de la nostra sang és superior al de la població mundial). Davant aquesta demostració, podem pensar que la màquina més espectacular fabricada per l'home és també una joguina maldestra.
La majoria dels fenòmens que es produeixen dins de la cèl·lula són de moment misteris. No obstant això, s'han aconseguit alguns avanços i el camí cap a la recerca està obert.
Fins fa pocs anys, les reaccions s'analitzaven de manera individual o col·lectiva en els tubs de cristall després de la dissolució de les cèl·lules. Aquesta estratègia ha suposat grans avanços. Entre altres coses, s'ha aconseguit conèixer l'estructura i origen, origen i destinació de molts compostos cel·lulars. Això ha permès aclarir els principals processos metabòlics com la glicolisis, el cicle de Krebs, etc. No obstant això, aquesta recerca, realitzada fora de l'arquitectura cel·lular, ha posat en enfosquiment molts altres detalls com la velocitat real, la ubicació i la direcció espacial dels processos que es produeixen a l'interior de la cèl·lula. Per exemple, les relacions entre les neurones a l'interior del cervell no s'han obtingut dades precises, ja que en un espai molt reduït es produeixen moltes i complexes reaccions.
Amb la finalitat de superar els obstacles esmentats anteriorment, els científics han tractat de desenvolupar noves tècniques que permetin analitzar aquests processos de la vida mantenint la integritat cel·lular. Aquestes noves tècniques es basen en l'espectroscòpia. És a dir, una vegada que una cèl·lula completa s'ha irradiat per ones electromagnètiques, les ones rebudes travessant la cèl·lula o per reflexió emmagatzemen informació sobre l'estructura cel·lular. En això consisteix la tècnica utilitzada en les radiografies. Els raigs X que obstrueixen els ossos no arriben a la pel·lícula, la qual cosa ens permet obtenir informació sobre la forma de l'os.
L'arquitectura molecular dins de la cèl·lula és molt complexa, però està composta per pocs tipus de molècules. Per exemple, totes les proteïnes d'un bacteri (unes 3.000) estan compostes per una combinació d'uns 20 aminoàcids diferents. Descobreix la fascinant habilitat de la vida. El mateix ocorre entre lípids i hidrats de carboni. El misteri està en la combinació!
Per tant, i desgraciadament davant les ones electromagnètiques, totes les proteïnes són similars, per la qual cosa no és possible analitzar el treball individual de cadascuna d'elles. No és aquest l'únic problema: d'una banda, la intensitat de la radiació utilitzada i la longitud d'ona (energia) han d'estar dins de límits que no afecten la cèl·lula. D'altra banda, les cèl·lules estan formades en gran manera per aigua i molts tipus d'ones no poden utilitzar-se perquè interfereixen amb l'aigua.
Si la recerca cel·lular ha hagut de fer front a molts problemes, alguna cosa semblança ha ocorregut quan s'ha tractat d'investigar l'espai exterior, en molts casos s'han utilitzat sondes. En altres paraules, s'han realitzat les màquines adequades i posteriorment s'han enviat a l'espai per a mesurar el paràmetre que es pretén investigar. El senyal de mesura la rebem per radi en la Terra.
En els treballs interns de les cèl·lules, fins i tot davant els problemes esmentats anteriorment, s'han creat molècules indicadores (o sondes). La sonda molecular ha de ser capaç de fer el mateix treball que la sonda espacial dins de la cèl·lula. Bàsicament les característiques principals d'aquesta mena de sondes són:
- El senyal espectroscòpic (resposta a les ones) de totes les molècules presents a l'interior de la cèl·lula és totalment diferent i totalment separada.
- Ha de ser un indicador concret d'una reacció o molècula perquè l'investigador pugui investigar els fenòmens individualment.
- El viatge a l'interior de la cèl·lula ha de realitzar-se sense provocar fatiga cel·lular. A més, la sonda ha de tenir una ubicació específica dins de la cèl·lula i coneguda: nucli, citoplasma, mitocondri, etc.
- L'estructura interna i la funció de la cèl·lula han de ser normals en el moment del mesurament. Evidentment, la sonda que aglutina totes aquestes
característiques no és fàcil de crear; totes les característiques de les màquines que enviem a l'espai han de ser incorporades dins d'una molècula. Afortunadament, la col·laboració entre químics orgànics i bioquímicos també ens ha donat resultats espectaculars en aquest camp. La fluorescència ha estat la seva eina més útil.
Què és la fluorescència?
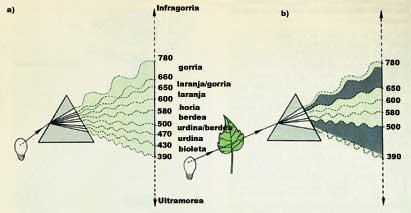
És un fenomen curiós que expliquen algunes molècules i que ha resultat molt útil. Com la llum està formada per ones, totes les molècules absorbeixen la llum d'una determinada longitud d'ona, travessen la resta d'ones o es reflecteixen a la volta. Com sabem, la clorofil·la és verda; si prenem com a exemple, veurem que irradiant amb llum blanca absorbeix la llum blava (que té una longitud d'ona entre 390 i 500 nm) i la vermella (que té una longitud d'ona entre 650 i 780 nm) i que els altres es veuen reflectits a la volta. El color de la llum reflectida és la diferència entre la llum radiada blanca i l'absorbida: llum verda (entre 500 i 650 nm). Per tant, totes les coses que tenen color lleven alguna cosa a la llum blanca. Els blancs res. Els negres tot. (Veure figura 1).
Què ocorre amb les ones de llum absorbides? Les ones de la llum, com qualsevol ona, transmeten energia i les molècules que absorbeixen les ones passen a l'estat excitat per l'energia rebuda. Perquè torni al seu estat energètic normal, emet aquesta energia d'una altra manera. En molts casos aquesta energia emesa és la calor formada per ones de baixa energia. En casos especials, no obstant això, l'energia d'excitació absorbida s'emet a través d'una altra ona visible i l'energia de la llum emesa és sempre inferior a la llum absorbida. Dit d'una altra manera, la molècula excitada expulsa la llum d'un altre color. I aquesta és precisament la fluorescència.
Les llums negres d'ús freqüent en les discoteques expulsen les ones d'alta energia que nosaltres no podem veure. No obstant això, aquestes ones tenen la capacitat d'excitar diverses molècules. Aquestes molècules excitades emeten, al seu torn, una llum blava-blanquinosa. Per exemple, les camises blanques, les dents i la tònica apareixen com si tinguessin llum pròpia (per efecte de la fluorescència).
Els compostos fluorescents naturals i artificials són coneguts i poden proporcionar informació precisa sobre els processos que es produeixen dins de la cèl·lula quan s'utilitzen com sonda.
Els indicadors fluorescents s'han utilitzat per a mesurar el pH intern de la cèl·lula. Els investigadors han col·locat a SNARF i fluorescent, grups que uneixen protons (H+), en una molècula fluorescent. A mesura que la molècula s'omple de protons, les seves característiques de fluorescència canvien i aquest fenomen és mesurable.
D'altra banda, l'ompliment de protons depèn de la concentració d'aquests. Per tant, com es pot apreciar en la figura, un canvi en la concentració dels protons implica la modificació de les característiques de fluorescència de l'indicador, que també pot mesurar-se amb gran precisió.
Mitjançant aquest sistema expert s'han hagut d'introduir aquest tipus de fluoróforos especials per a mesurar el pH intern de la cèl·lula sense provocar fatiga cel·lular. No obstant això, el sistema no pot posar-se a treballar de qualsevol manera. Queden alguns problemes tècnics que caldrà superar i no de qualsevol tipus. Els grups que uneixen protons tenen càrrega elèctrica i interactuen amb l'aigua (són per tant hidròfils).
D'altra banda, la cèl·lula té una membrana cel·lular formada per proteïnes i lípids en la vora exterior, que controla bé el trànsit molecular entre l'exterior i l'interior. Aquest porter molecular no li permet introduir-se en qualsevol molècula carregada que no coneix i en tractar-se d'un indicador fluorescent artificial, no troba porta d'entrada. (Veure 2. imatge)
Davant aquest problema, els científics han aconseguit disfressar als grups carregats mitjançant l'esterificació. Amb aquest canvi químic s'aconsegueix passar d'una molècula hidròfila a una hidròfoba, la qual cosa li permet travessar fàcilment la barrera lipídica que té davant sense passar per la porta especial. Una vegada dins, a causa de l'enzim esterasa que conté les cèl·lules, l'èster hidrofòbic recupera l'estructura original i el fluoroforo actiu pot actuar com a indicador dins de la cèl·lula.

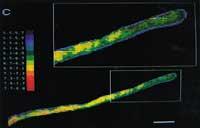
En alguns casos, l'indicador fluorescent queda en el citosol, però en uns altres s'acumula en parts especials de l'estructura interna de la cèl·lula. Un sol canvi en el grup químic pot tenir gran influència en l'emplaçament cel·lular de la molècula. Per exemple, la fluoresceïna queda en el citoplasma, però la carboxifluoresq s'acumula especialment en el vaquer de les cèl·lules vegetals. Aquestes diferències s'han utilitzat per a la realització d'estudis especials de fragments cel·lulars (veure figura 3).
Els mesuraments de pH de la cèl·lula han donat resultats molt importants, però el treball que més fama ha donat als indicadors fluorescents és, sens dubte, el mesurament de la concentració de calci(II) dins de la cèl·lula. Les recerques dutes a terme en els últims anys han demostrat que aquest simple catió té una gran responsabilitat en el comportament intern de la cèl·lula, sent la seva comesa el del segon missatger.
Normalment, la concentració de calci(II) de la cèl·lula (en el llenguatge dels científics [Ca 2+ ]) és molt baixa en el citoplasma (entorn de 10-7 M) pel fet que la major part de Ca 2+ de la cèl·lula es concentra en compartiments especials. S'acumula principalment en animals en reticle endoplasmàtic i plantes en vacúols.
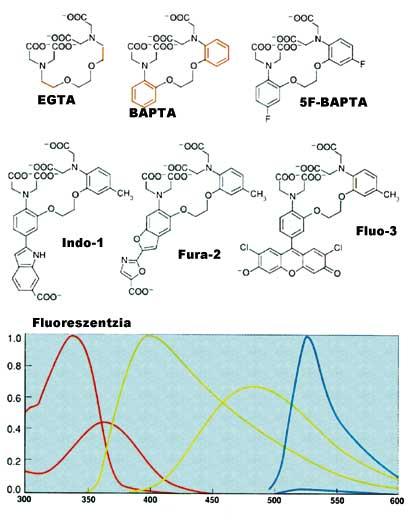
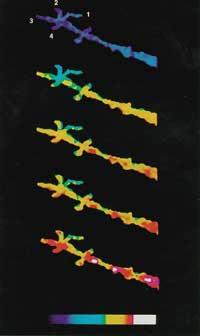
Després de rebre certs senyals o estímuls externs, el calci emmagatzemat en els compartiments és expulsat i el citoplasma ascendeix fins a 10 -5 M. Aquest augment té una gran influència en les proteïnes especials i genera cadenes de reacció. En l'extrem final d'aquestes cadenes de reacció es troba l'expressió dels gens. Per tant, és molt important poder mesurar els canvis de calci(II) dins de la cèl·lula. El primer indicador fluorescent per a mesurar Ca 2+ va ser creat per Roger Tsien. EGTA, a partir del conegut complex càlcic, crea la nova estructura BAPTA. Gestió de residus Aquest canvi modifica les característiques espectrals de BAPTA. Desgraciadament, l'espectre d'absorció del BAPTA està pròxim a la resta dels compostos que conté la cèl·lula, la qual cosa comporta problemes de mesurament. Per això, Tsien va col·locar altres grups fluorescents mantenint el caràcter molecular i les característiques del BAPTA, formant Quin-2, Fura-2, Indo-1 i finalment Fluo-3 (veure figura 4).
Actualment es pot mesurar a tota velocitat [Ca 2+ ] de la cèl·lula. Aquestes tècniques han demostrat que les respostes químiques en les cèl·lules són molt ràpides i precises (veure figura 5).
L'última revolució: veure les proteïnes
Les proteïnes són un dels actors més representatius de les cèl·lules. La informació codificada en els gens s'expressa en proteïnes i aquestes són les molècules treballadores que realitzen les diferències. Per tant, veure les diferents proteïnes treballant dins de la cèl·lula ha estat un dels principals objectius que els científics han seguit sense cansar-se. Gràcies a la col·laboració entre l'enginyeria genètica i la bioquímica s'han publicat espectaculars resultats en els últims anys.

Dins del codi d'informació que té cada proteïna es pot trobar el senyal de la seva adreça. Aquest senyal es troba en l'extrem anterior de la proteïna i li obre totes les portes al lloc que anirà. Igual que la carta enviada per correu, la cèl·lula conduiria un complex procés de distribució de cada proteïna. Finalment, una vegada aconseguida la proteïna en el seu emplaçament, el senyal de direcció s'elimina i en alguns casos se separa d'altres àrees funcionals. En altres ocasions continua formant part de la proteïna.
Alguns animals marins contenen proteïnes que per si mateixes són fluorescents. A causa d'aquestes proteïnes especials, les meduses són conegudes des de fa temps i han estat utilitzades com a indicadors fluorescents per a aclarir la direcció d'altres proteïnes.
Les bases d'aquesta tècnica espectacular són senzilles: es pot aïllar prèviament el gen de la proteïna que es vol investigar i col·locar el gen de l'indicador fluorescent darrere o en el centre del gen; posteriorment s'introdueix una nova còpia a l'interior de la cèl·lula perquè continuï el seu camí. En absència d'artefactes, l'indicador fluorescent permet obtenir una imatge exacta i espectacular de quan, on i quant fa la proteïna investigada.
A la vista de les innovacions existents, es pot dir que amb l'ús de cèl·lules vives es publicaran molts descobriments en el futur; la bellesa dels resultats obtinguts ens aportarà molt més que el plaer del coneixement.
Zu idazle
Zientzia aldizkaria

