

El niu de Kukua: una competició contínua
Per a la ciència, per contra, han estat desconegudes algunes de les característiques del contingent, mentre que unes altres han estat repletes d'especulació, sovint a causa de les conviccions populars esmentades. Veurem, per tant, la naturalesa d'aquest ocell tan popular i, al mateix temps, poc coneguda.
Paràsit niu

Tot ésser viu ha d'afrontar la reproducció per a completar el seu cicle de vida i la complexitat de les seves estratègies reproductives en la naturalesa és enorme. Un dels més sorprenents és el niu paràsit. Des que Aristòtil, fa 2.300 anys, descrivís la tendència paràsita del contingent, s'ha investigat molt de sobre aquest tema i avui dia es coneix molt bé el mecanisme d'aquest comportament. No obstant això, els factors que han contribuït a aquesta estratègia al llarg de l'evolució, l'eficàcia i les causes d'aquest comportament, no són prou estudiats, presentant llacunes significatives des del punt de vista de l'ecologia evolutiva.
El principal problema de molts éssers són altres vius (depredadors, paràsits, etc.) i han evolucionat per a afrontar-los. El depredador també ha evolucionat, entrant en una contínua competència armamentística. Així, moltes característiques són conseqüència directa d'una coevolución, és a dir, adaptacions que s'han desenvolupat per a superar els obstacles i dificultats que un grup d'éssers, un altre, li ha posat. És en aquest context on hem de situar la confrontació evolutiva que ha tingut i té el bufó i els seus hostalers. D'una banda, tenim les estratègies que el bufó ha utilitzat per a enganyar a l'hostaler i, per un altre, les adaptacions que els hostalers han desenvolupat per a fer front al frau del contingent.

Les kuku femelles deixen els ous mimètics (molt similars en grandària, color i forma a la de l'hoste) en el niu d'alguna altra espècie, i són elles les encarregades de la seva elaboració. A pesar que molta gent ha considerat que l'únic hostaler del contingent és el txantxangorri, en tota Europa es coneixen més de 100 hostalers; a Euskal Herria, els hostalers més comuns que sofreixen aquest descarat passerell són: Txepetxa ( Troglodytes troglodytes ), pit-roig ( Erithacus rubecula ), carricero comú ( Acrocephalus scirpaceus ), carracín de carrichaca ( Acrocephalus arundinaceus), terrisseria blanca ( Motacilla alba ), tatenálama ( Tochantus). L'elecció dels hostalers no és aleatòria; tots són 2-3 vegades més petits que el bufó i són insectívors. La mesura de l'hoste, com veurem més endavant, està relacionada amb els ous de la cuba i per descomptat la presència d'insectívors amb l'alimentació del txito de bufó.
En la selecció d'hostalers, el desenvolupament del chitaje és de vital importància. El bufó sempre anirà al niu després de la posada de l'hoste. Però si l'allotjament està molt avançat, trenca el niu obligant l'hoste a construir un niu nou i a posar més ous. De fet, en els chites més avançats, els pollets de l'hoste naixerien abans i en néixer el bufó els altres estarien fenedures; en aquesta situació, el cuquecillo no necessitaria aliment degut a la competència dels niu.

Per tant, l'època de la compassió del contingent està condicionada pel chitazo dels hostalers. Els bufons petits solen estar vigilats, cuiden als hostalers i sovint observen com construeixen el niu. Si els hostalers s'adonen d'això, ataquen el contingent. Les femelles de bufó aprofiten l'absència dels nidificants per a fresar l'ou, podent realitzar aquesta acció en deu segons. Es posen cada dos dies, normalment un ou per niu i de 8 a 15 ous cadascun. Abans de dipositar l'ou en el niu de l'hoste, roben un ou de l'hoste, igualant així el niu.
Aquest robatori d'ous ha generat falses conviccions. S'ha pensat que el bufó posa els ous prop del niu de l'hoste, els pren en el pic i els deixa dins del niu. Aquesta creença s'ha utilitzat per a explicar els paràsits que es produeixen en els nius d'entrada estreta o complicada. El bufó realitza sempre directament sobre el niu de l'hoste i per a això disposa de diverses adaptacions: potes llargues i fortes, per a subjectar i col·locar el niu de l'hoste, la claveguera també és allargada i la pela gruixuda de l'ou, en els nius tancats on el bufó no entra bé, els ous de posada superior no es trenquen fàcilment.
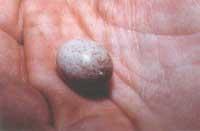
Cada kuku femella només pot posar un tipus d'ou, per la qual cosa només tindrà l'hoste corresponent. En conseqüència, trobarem diferents llinatges de femella basats en la forma i el color dels ous. D'aquesta manera, una femella cucut posa els ous en forma d'hoste que la va criar. Per exemple, si un bufó ha nascut en el niu de la txantxangorri, aquest també pondrà com la txantxangorri i no podrà pondre en el niu de la txepetxea, ja que això ho reconeixeria. A través dels experiments, encara que això està demostrat, no s'investiguen les conseqüències genètiques d'aquest fenomen.
Es creu que el mecanisme genètic que condiciona les característiques dels ous només el porta la femella, per la qual cosa els mascles no produeixen canvis en les característiques de l'ou en l'encreuament: existeixen llinatges de femella diferents i els mascles garanteixen la unitat de l'espècie.

En general, els hostalers no admeten ous estranys en els nius i expulsen als que són diferents dels seus ous. Aquesta discriminació dels hostalers ha estat la pressió evolutiva que ha impulsat els ous mimètics, ja que només els d'aspecte dels ous de l'hoste podran enganyar a l'hoste. En qualsevol cas, aquesta no ha estat l'única raó, perquè la competència entre bufons també ha afavorit aquest mimetisme. Si a un niu amb ou de bufó s'aproxima un segon bufó, el més rendible serà, per descomptat, l'expulsió de l'ou del bufó i no de l'hoste. El primer txito naixerà de l'ou del primer bufó, que eliminarà tots els ous que trobi.
El mimetisme de l'ou es basa en dues característiques: el color i la talla. En general, el color sol ser molt semblant a l'ou de l'hoste, segons el cas, amb el seu propi pintar. Però el mimetisme tampoc es dóna en tots els casos. Aquestes excepcions poden ser molt significatives i tenim un bon exemple, el cas del tuntún. El tuntún pon blaus i el bufó que el parasita és blanquinós. Per tant, si l'ou mimètic ha afavorit la discriminació dels hostalers, com explicar aquesta falta de mimetisme?

En diferents estudis realitzats amb tuntu (en el niu amb ous de diferents colors i formes) s'ha demostrat que el tuntún admet tots els ous de qualsevol color i grandària. I no, el tuntu no és cec, però sembla que el tuntún és un nou hoste del bufó i que encara no “aprèn” a rebutjar els ous. Per a confirmar això, es pot realitzar un altre tipus de recerca: observar com es comporten davant l'ou de la cuba, alguns txokos, encara que no són els hostalers del contingent. De fet, alguns ocells que no són hostals mostren un major grau de discriminació que l'hostal comú.
Per exemple, l'ocell mosca grisa ( Muscicapa striata ), en posar l'ou de la cuba, el separa amb facilitat, i en l'actualitat no s'inclou en la llista d'hostes del contingent. Seguint el mateix criteri que hem utilitzat per a explicar el fenomen del tuntún, podem dir que l'ocell mosca grisa, en el seu moment, va ser paràsit del bufó i que en la competició havia superat el bufó.
L'ou del bufó és molt petit en relació al seu cos (proporcionalment tres vegades menor que el de la resta d'ocells), sent una mica major que el de l'hoste. En aquest cas també hi ha raons de mimetisme, però hi ha unes altres: el bufó produeix un gran nombre d'ous (8-15), la qual cosa suposa una despesa considerable. Per això, gràcies als balanços energètics, la producció d'ous petits ha permès augmentar el seu número. I quants més ous posis, més cries, clar.

El kukukume naixerà després del chitaje de 11-12 dies, abans que els nidificants (la majoria dels hostalers tenen un chitaje de 14-15 dies) i monopolitzarà l'atenció i el menjar dels pares. Sembla ser que la incubació s'inicia en l'obsequi de la galleda, i aquesta és la raó per la qual naixia abans en el niu. Aquest avançament és de vital importància, ja que el cucachorro tindrà l'oportunitat de treure els ous de l'hoste, eliminant així als competidors.
Són pollastres nounats, silenciosos i cecs, però no triguen a empènyer els ous que té al seu costat, posant-los al capdavant, estrenyent les potes fortes contra les parets del niu i posant l'ou a l'esquena, llancen l'ou del niu. En 3-4 hores llançarà tots els ous. Moltes vegades els pares són testimonis, però no fan cap gest per a evitar el desastre. En alguns casos, el cucachorro, més que amb ous, es troba en el niu amb pollastres nounats i en aquests casos s'ofega.
Quan s'ha convertit en amo del niu, tots els insectes portats pels pares seran per a ell i en pocs dies superarà la seva grandària. Es converteix en un espectacle únic, pujant la txepetxa sobre el cap del kukukume i descobrint com li ofereix el menjar, orgullós de la seva suposada cria.

A les 4-5 setmanes, el chito es va acostant a les branques pròximes, amb vols curts, encara que s'alimentin dels aliments que els porten els pares. Una o dues setmanes després abandona completament el niu i comença a preparar el llarg viatge a Àfrica, on ingerirà gran quantitat de greix. Per al segon any són capaços de reproduir els bufons.
El mimetisme dels ous és molt precís, mentre que el kukukume no s'assembla als chitos dels hostalers. Per què es mostren tan prudents a l'hora d'acceptar ous estranys i, per contra, estan disposats a criar qualsevol pollet, encara que sigui totalment diferent i molt més gran que ells?
Podria pensar-se que l'hoste separa l'ou del bufó, comparant-lo amb els seus, i amb els chitos, no el pot fer perquè només està el cuquero. Però això no és així, ja que s'ha vist que els chites mixtos (els chitos del bufó i de l'hoste) creixen sense problemes. Per tant, al no existir discriminació respecte al chito, la selecció natural no ha impulsat el txito mimètic.
Encara que no tots els autors estan d'acord, sembla que la principal causa d'aquesta falta de discriminació és la rendibilitat típica de la naturalesa: el desenvolupament d'un mecanisme de separació dels ous estranys per part de la selecció natural resulta avantatjós, ja que l'hoste és a temps d'expulsar l'ou i de salvar la incubació; o com en altres casos, abandonar el niu i construir un nou per a tirar endavant el txitaldia. Per contra, el desenvolupament de mecanismes per a aprendre a diferenciar el chito de bufó pot servir per a molt poc, ja que ja és tard per a iniciar un nou chitazo. Per això, els hostalers no incuben cap ou, però el primer chito que veuen davant sol menjar per si de cas.
Malgrat el risc de marejar-se en el túnel del temps en analitzar l'evolució, moltes vegades és imprescindible submergir-se en ell, ja que alguns comportaments i adaptacions només així es poden entendre. I a més, com hem vist, no hem d'anar molt lluny a analitzar comportaments estranys, perquè hem d'investigar les causes de moltes conductes que ocorren en el nostre entorn.

Document d'identitat de Kukua
El bufó ( Cuculus canorus ) és com un colom modal. Cua molt llarga d'uns 13-15 cm. Cap relativament petit i ales afilades. Els adults solen tenir cap, pit i suport gris-blaus. El ventre és blanquinós, amb línies transversals marrons i estretes. Les ales i la cua són les parts més fosques del cos, ja que són gairebé negres. La cua està decorada amb tipus blancs. El mascle i la femella són molt similars i ocasionalment es poden trobar cucles femelles molt marrons.
Els juvenils són vermellosos o grisos. Quan vola s'assembla molt a l'esparver ( Accipiter nisus ). En el sòl té una maldestra marxa i quan canta canvia la silueta habitual, ja que baixa el cap, tanca les ales, aixeca la cua i la fa molt ampla. El famós cant és el de la falla (kuku), que té un cant més baix, més baix i més desconegut.

El contingent té una àmplia distribució geogràfica. Incuba gairebé per tota Europa i Àsia, excepte en tundra i deserts. Hiverna en el sud d'Àfrica. La seva distribució geogràfica a Àfrica no és molt coneguda, però sembla que la majoria es concentra a Namíbia i Sud-àfrica. A Euskal Herria tenim una espècie típica en l'època del Txitaldi.
Quant al seu lloc de residència no presenta grans exigències i apareix pràcticament a tot arreu, excepte en muntanyes nues, zones forestals molt extenses, illes oceàniques i zones molt urbanitzades. No obstant això, està condicionada pel valor principal que requereix l'establiment de l'hàbitat, és a dir, per l'ús hostaler. A l'hora de menjar descendeixen als prats, però no els agraden els prats molt extensos, per la qual cosa els hàbitats més comuns són els boscos intercalats amb els simples.
Com hem esmentat, el bufó és un ocell migrador que hiverna a Àfrica. Això va obrir la creença que el bufó es convertia en gavina a l'hivern, com deia Aristòtil: “kokkus metaballei ex ierakos” i aquesta creença encara es manté en diverses localitats. D'altra banda, amb motiu d'aquesta migració anual, Azkue, en l'antic basc, deia que, “una vegada a l'any”, es deia “de bufó a bufó”.
El viatge al Sud s'inicia en les proximitats d'Agost i els adults arriben abans que els joves. A Àfrica romanen fins a Abril. A la primavera arriben fins a Abril-Maig, primer mascles i després femelles i joves d'un any. Aquest llarg viatge el realitzen de nit i en petits grups. Els joves d'un any, en general, tornen als voltants del lloc on van néixer i els adults també tenen una gran fidelitat a les zones de chiteo anuals.
Li agraden els escarabats i les erugues per a menjar, inclosos els peluts i verinosos dels altres ocells. També pot menjar aranyes, cucs, melses, rosegadors, granotes i raps joves, ous d'ocells i chikis. Encara que la majoria de les vegades busquen farratge, solitari i col·locat en els miradors, en el sòl també s'ha vist agitant les plantes amb les ales, espantant els insectes i obtenint l'aliment amb facilitat. Abans de la seva ingestió, dota a les herbes peludes i verinoses de fortes sacsejades perquè llancin el contingut intestinal. Per a fer front als verinosos pèls de les erugues, tenen el paparo modelat, ja que de tant en tant els teixits mucosos del lloc es muden i els pèls són expulsats al vòmit.
L'elecció dels reproductors i la seva relació no es coneix molt bé, però sembla que les femelles s'equipessin cada any amb més d'un mascle i a més cada any amb diversos mascle. No obstant això, atès que el ràtio sexual (proporció ar:femella) de la majoria de les poblacions analitzades és de 1:1, no es descarta l'existència d'una altra mena d'equiparació. A la primavera, quan els mascles arriben a les zones de chitaje, es fixen els territoris mitjançant cançons. Després, amb l'arribada de les femelles, les cançons es van esmorteint; més endavant, quan s'acosta l'època de la raqueta, tornem a cantar. Finalment, a la fi d'estiu es tallen les cançons, encara que encara hi ha dies en què el bufó es mou entre nosaltres.
Zu idazle
Zientzia aldizkaria

