

Las bacterias también tienen esqueleto
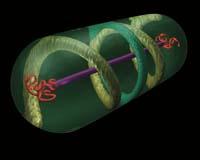
Perritos confusos. Hasta hace poco las bacterias eran en los ojos de los biólogos: sacos que contienen material genético, algunas enzimas y otras moléculas mezcladas; sin estructuras estables, sin orden, sin esqueletos. Pero las cosas están cambiando. Dentro de estos sacos se han encontrado diversas estructuras. Muchos de ellos equivalentes a los componentes del citoesqueleto de células eucariotas.
En las células eucariotas, es decir, de plantas, animales, hongos y protistas, es muy conocido el entramado interior denominado citoesqueleto. Es una estructura dinámica, compuesta por tres tipos de componentes: microfilamentos (actina), filamentos medios y microtúbulos (tubulina). Y cumple varias funciones: dar y mantener forma a las células, facilitar el movimiento, transporte intracelular y división celular...
Se consideraba que las bacterias no tenían citoesqueleto, por lo que se consideraba que era el desarrollado tras la formación de eucariotas (procedentes de bacterias). Pero ahora parece que no fue así: el origen de las actinas, filamentos medios y tubulinas podría estar en bacterias.
Las bacterias han pasado de ser protagonistas secundarios a estar de moda en el campo de la biología celular, y los descubrimientos se suceden.
Detrás de las formas

Se puede decir que en 2001 la rueda de los descubrimientos tomó el ritmo. En ese año, un grupo de investigadores de la Universidad de Newcastle en Inglaterra consiguió por primera vez ver la estructura de la proteína MreB dentro de la bacteria Bacillus subtilis: Las fibras MreB forman una hélice bajo la pared de las bacterias cilíndricas.
La proteína MreB es muy parecida a la actina de los eucariotas y puede tener el mismo origen. Además, cumplen una función similar. En las bacterias cilíndricas las proteínas MreB y muy próximas a ella tienen una función estructural, es decir, dan forma a la bacteria.
De hecho, los investigadores han visto que si les falta esta proteína cambia su aspecto. B. subtilis, el MreB afecta a la anchura de la bacteria y el Mbl a la longitud; y E. Las bacterias coli pasan de ser cilíndricas a esféricas cuando se les quita el MreB.
De hecho, las bacterias esféricas no tienen MreB. Más aún, analizando los genomas de diversas bacterias y arqueos (los dos grupos de procariotas), los investigadores de Newcastle han descubierto que los procariotas cilíndricos, filamentosos y helicoidales contienen uno o más genes de la proteína MreB, mientras que los esféricos tienen ninguno.
Por otra parte, en formas más complejas de ciertas bacterias es muy probable que el citoesqueleto esté relacionado. De momento, sólo han encontrado un ejemplo: Las bacterias Caulobacter crescentus tienen forma de alubias, y parece que la otra proteína les da esa curvatura. Al retirar esta proteína estas bacterias adquieren forma cilíndrica. Esta proteína es krescentina y es muy parecida al filamento medio de los eucariotas.
Claves de la división
Al margen de las formas, el citoesqueleto también tiene gran importancia en la división celular. Y en eso los procariotas no tienen nada en común. Las bacterias presentes en el medio apropiado pueden dividirse cada media hora. Pues una de las claves de esta eficacia puede estar precisamente en el citoesqueleto. Pero los científicos no han hecho más que empezar a aclarar los mecanismos.
La proteína FtsZ es uno de los protagonistas de la división celular de los procariotas. Forma un anillo en la zona donde se realiza la división, en el centro de la célula. Al retirar esta proteína a los bacilos cilíndricos, se estiran y estiran sin dividirse en dos. Por su parte, los Staphylococcus aureus esféricos, una vez eliminado el gen de FtsZ, comienzan a construir la nueva pared celular en cualquier lugar y pueden llegar a aumentar el volumen ocho veces antes de que explote.
De alguna manera, la proteína FtsZ controla dónde hay que construir la nueva pared. A su alrededor recoge las proteínas que construyen la pared. En el caso de S. aureus forma el anillo en el ecuador y gobierna la creación de los dos nuevos hemisferios. En los bacilos, por su parte, parece que en un principio la proteína MreB provoca el estiramiento de la célula y luego entra en juego el FtsZ, creando nuevos extremos redondeados para dividir la célula en dos.
Los anillos de FtsZ pueden tener gran importancia en el origen de la división celular. Un grupo de investigadores de la Universidad Duke de Carolina del Norte ha visto recientemente que esta proteína también forma anillos dentro de las gotas microscópicas de aceite. Ahora, aunque la división celular de las bacterias es un mecanismo más complejo, los investigadores creen que las células más primitivas tendrían suficiente FtsZ para dividirse en dos.
Por otra parte, algunos orgánulos de células eucariotas, las mitocondrias y los cloroplastos utilizan el anillo de FtsZ para dividirse. Esto refuerza, en definitiva, la creencia de que estos orgánulos son procariotas que en su día fueron introducidos en una célula eucariota.
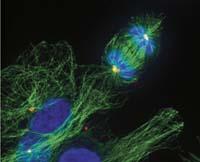
Herramientas modificadas
Los eucariotas también tienen una especie de proteína FtsZ, la tubulina. Y la tubulina también participa en la división celular, pero cumple una función muy diferente: mantiene alejados los cromosomas correspondientes a cada nueva célula durante la división. De esta forma se asegura que cada nueva célula tendrá sus cromosomas.
Es curioso que estas dos proteínas de un mismo origen cumplan una función tan diferente. Es decir, FtsZ y la tubulina son dos versiones de una misma herramienta que los procariotas y eucariotas utilizan para realizar trabajos muy diferentes. Para el mismo trabajo utilizan herramientas muy diferentes. Por un lado, para el trabajo de la proteína FtsZ, equivalente a la tubulina, los eucariotas utilizan una herramienta de actina. Y por otro lado, los procariotas hacen lo que los eucariotas hacen con la tubulina con una proteína equivalente a la actina.
Y es que, al igual que los eucariotas, las bacterias también deben distribuir bien el material genético en la fragmentación. Las bacterias poseen un único cromosoma, además de unos anillos de ADN llamados plasmidos. Los plásmidos aportan resistencia a los antibióticos y otros 'extra' a las bacterias, por lo que es muy importante que las nuevas bacterias se queden con una copia de los plásmidos.
Pues al igual que hacen las tubulinas con los cromosomas de los eucariotas, la proteína ParM produce la separación de algunos plasmidos, alejando las copias. Y ese ParM es una versión bacteriana de la actina.
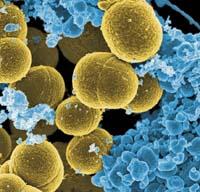
ParM es una cadena de proteínas con capacidad de autoadición y eliminación de unidades. La cadena ParM se une a las dos nuevas copias del plásmido a repartir. Posteriormente, mediante la adición de nuevas unidades se inicia el estiramiento de la cadena, que envía los dos plásmidos a los dos extremos de la célula.
Este es el mecanismo de distribución de algunos plasmidos. ¿Pero qué pasa con el cromosoma de bacterias? ¿Cómo distribuyen las copias del cromosoma? Eso sigue siendo un misterio.
Y no es la única. Queda un largo camino por recorrer para comprender bien la organización y funcionamiento de las células de los procariotas. Además de los ya mencionados, han encontrado muchos más elementos del citoesqueleto y se encuentran. También proteínas que no tienen nada en el citoesqueleto de los eucariotas. Pero poco sabemos de los mecanismos que están detrás de todos estos elementos. Todavía queda mucho por investigar dentro de estos perritos.
Zu idazle
Zientzia aldizkaria

