

Els bacteris també tenen esquelet
Gossets confusos. Fins fa poc els bacteris eren en els ulls dels biòlegs: sacs que contenen material genètic, alguns enzims i altres molècules barrejades; sense estructures estables, sense ordre, sense esquelets. Però les coses estan canviant. Dins d'aquests sacs s'han trobat diverses estructures. Molts d'ells equivalents als components del citoesquelet de cèl·lules eucariotes.
En les cèl·lules eucariotes, és a dir, de plantes, animals, fongs i protistos, és molt conegut l'entramat interior denominat citoesquelet. És una estructura dinàmica, composta per tres tipus de components: microfilaments (actina), filaments mitjans i microtúbuls (tubulina). I compleix diverses funcions: donar i mantenir forma a les cèl·lules, facilitar el moviment, transport intracel·lular i divisió cel·lular...
Es considerava que els bacteris no tenien citoesquelet, per la qual cosa es considerava que era el desenvolupat després de la formació d'eucariotes (procedents de bacteris). Però ara sembla que no va ser així: l'origen de les actines, filaments mitjans i tubulines podria estar en bacteris.
Els bacteris han passat de ser protagonistes secundaris a estar de moda en el camp de la biologia cel·lular, i els descobriments se succeeixen.
Darrere de les formes

Es pot dir que en 2001 la roda dels descobriments va prendre el ritme. En aquest any, un grup d'investigadors de la Universitat de Newcastle a Anglaterra va aconseguir per primera vegada veure l'estructura de la proteïna MreB dins del bacteri Bacillus subtilis: Les fibres MreB formen una hèlix sota la paret dels bacteris cilíndrics.
La proteïna MreB és molt semblant a l'actina dels eucariotes i pot tenir el mateix origen. A més, compleixen una funció similar. En els bacteris cilíndrics les proteïnes MreB i molt pròximes a ella tenen una funció estructural, és a dir, donen forma al bacteri.
De fet, els investigadors han vist que si els falta aquesta proteïna canvia el seu aspecte. B. subtilis, el MreB afecta a l'amplària del bacteri i el Mbl a la longitud; i E. Els bacteris coli passen de ser cilíndriques a esfèriques quan se'ls lleva el MreB.
De fet, els bacteris esfèrics no tenen MreB. Més encara, analitzant els genomes de diversos bacteris i arquejos (els dos grups de procariotes), els investigadors de Newcastle han descobert que els procariotes cilíndrics, filamentosos i helicoidals contenen un o més gens de la proteïna MreB, mentre que els esfèrics tenen cap.
D'altra banda, en formes més complexes de certs bacteris és molt probable que el citoesquelet estigui relacionat. De moment, només han trobat un exemple: Els bacteris Caulobacter crescentus tenen forma de mongetes, i sembla que l'altra proteïna els dóna aquesta curvatura. En retirar aquesta proteïna aquests bacteris adquireixen forma cilíndrica. Aquesta proteïna és krescentina i és molt semblant al filament mitjà dels eucariotes.
Claus de la divisió
Al marge de les formes, el citoesquelet també té gran importància en la divisió cel·lular. I en això els procariotes no tenen res en comú. Els bacteris presents en el mig apropiat poden dividir-se cada mitja hora. Perquè una de les claus d'aquesta eficàcia pot estar precisament en el citoesquelet. Però els científics no han fet més que començar a aclarir els mecanismes.
La proteïna FtsZ és un dels protagonistes de la divisió cel·lular dels procariotes. Forma un anell en la zona on es realitza la divisió, en el centre de la cèl·lula. En retirar aquesta proteïna als bacils cilíndrics, s'estiren i estiren sense dividir-se en dos. Per part seva, els Staphylococcus aureus esfèrics, una vegada eliminat el gen de FtsZ, comencen a construir la nova paret cel·lular en qualsevol lloc i poden arribar a augmentar el volum vuit vegades abans que exploti.
D'alguna manera, la proteïna FtsZ controla on cal construir la nova paret. Al seu al voltant recull les proteïnes que construeixen la paret. En el cas de S. aureus forma l'anell en l'equador i governa la creació dels dos nous hemisferis. En els bacils, per part seva, sembla que al principi la proteïna MreB provoca l'estirament de la cèl·lula i després entra en joc el FtsZ, creant nous extrems arrodonits per a dividir la cèl·lula en dues.
Els anells de FtsZ poden tenir gran importància en l'origen de la divisió cel·lular. Un grup d'investigadors de la Universitat Duke de Carolina del Nord ha vist recentment que aquesta proteïna també forma anells dins de les gotes microscòpiques d'oli. Ara, encara que la divisió cel·lular dels bacteris és un mecanisme més complex, els investigadors creuen que les cèl·lules més primitives tindrien suficient FtsZ per a dividir-se en dues.
D'altra banda, alguns orgànuls de cèl·lules eucariotes, els mitocondris i els cloroplastos utilitzen l'anell de FtsZ per a dividir-se. Això reforça, en definitiva, la creença que aquests orgànuls són procariotes que en el seu moment van ser introduïts en una cèl·lula eucariota.
Eines modificades
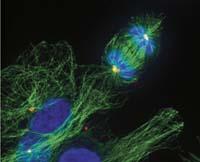
Els eucariotes també tenen una espècie de proteïna FtsZ, la tubulina. I la tubulina també participa en la divisió cel·lular, però compleix una funció molt diferent: manté allunyats els cromosomes corresponents a cada nova cèl·lula durant la divisió. D'aquesta forma s'assegura que cada nova cèl·lula tindrà els seus cromosomes.
És curiós que aquestes dues proteïnes d'un mateix origen compleixin una funció tan diferent. És a dir, FtsZ i la tubulina són dues versions d'una mateixa eina que els procariotes i eucariotes utilitzen per a fer treballs molt diferents. Per al mateix treball utilitzen eines molt diferents. D'una banda, per al treball de la proteïna FtsZ, equivalent a la tubulina, els eucariotes utilitzen una eina d'actina. I d'altra banda, els procariotes fan el que els eucariotes fan amb la tubulina amb una proteïna equivalent a l'actina.
I és que, igual que els eucariotes, els bacteris també han de distribuir bé el material genètic en la fragmentació. Els bacteris posseeixen un únic cromosoma, a més d'uns anells d'ADN anomenats plasmidos. Els plasmidis aporten resistència als antibiòtics i altres 'extra' als bacteris, per la qual cosa és molt important que els nous bacteris es quedin amb una còpia dels plasmidis.
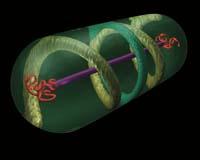
Perquè igual que fan les tubulines amb els cromosomes dels eucariotes, la proteïna ParM produeix la separació d'alguns plasmidos, allunyant les còpies. I aquest ParM és una versió bacteriana de l'actina.
ParM és una cadena de proteïnes amb capacitat d'autoadición i eliminació d'unitats. La cadena ParM s'uneix a les dues noves còpies del plasmidi a repartir. Posteriorment, mitjançant l'addició de noves unitats s'inicia l'estirament de la cadena, que envia els dos plasmidis als dos extrems de la cèl·lula.
Aquest és el mecanisme de distribució d'alguns plasmidos. Però què passa amb el cromosoma de bacteris? Com distribueixen les còpies del cromosoma? Això continua sent un misteri.
I no és l'única. Queda un llarg camí per recórrer per a comprendre bé l'organització i funcionament de les cèl·lules dels procariotes. A més dels ja esmentats, han trobat molts més elements del citoesquelet i es troben. També proteïnes que no tenen res en el citoesquelet dels eucariotes. Però poc sabem dels mecanismes que estan darrere de tots aquests elements. Encara queda molt per investigar dins d'aquests gossets.
Zu idazle
Zientzia aldizkaria

