

Non hegoa, là la jambe

Cela a été l'objectif de cette thèse de doctorat que nous allons vous dire: déterminer la niche écologique d'un animal inconnu qui a été jusqu'à récemment hors de l'empreinte des scientifiques. Je veux dire le grand chauve-souris alpin (Plecotus macrobullaris), un animal très curieux et intéressant, comme vous le verrez. Il a été décrit pour la première fois en 2003 dans les Alpes autrichiennes et pendant quatre ans il a été responsable de mes sueurs, casseroles et jouissance.
En 2010, déjà si lointain, je me suis demandé pourquoi cette chauve-souris, contrairement au reste, est liée aux hautes montagnes. À la recherche d’une réponse, j’ai commencé à analyser la niche écologique de l’herbacé alpin avec les membres du groupe de travail Écologie et Évolution du Comportement de l’UPV/EHU: Où est enregistré cet animal? Où chassez-vous ? Quelles températures pouvez-vous supporter? Que mangez-vous ? Chacune de ces questions fait référence à un axe de niche écologique, et ce que je vais raconter ci-dessous est ce que j'ai appris dans les quatre années que nous avons passées à chercher des réponses.
Niche écologique grinnellois
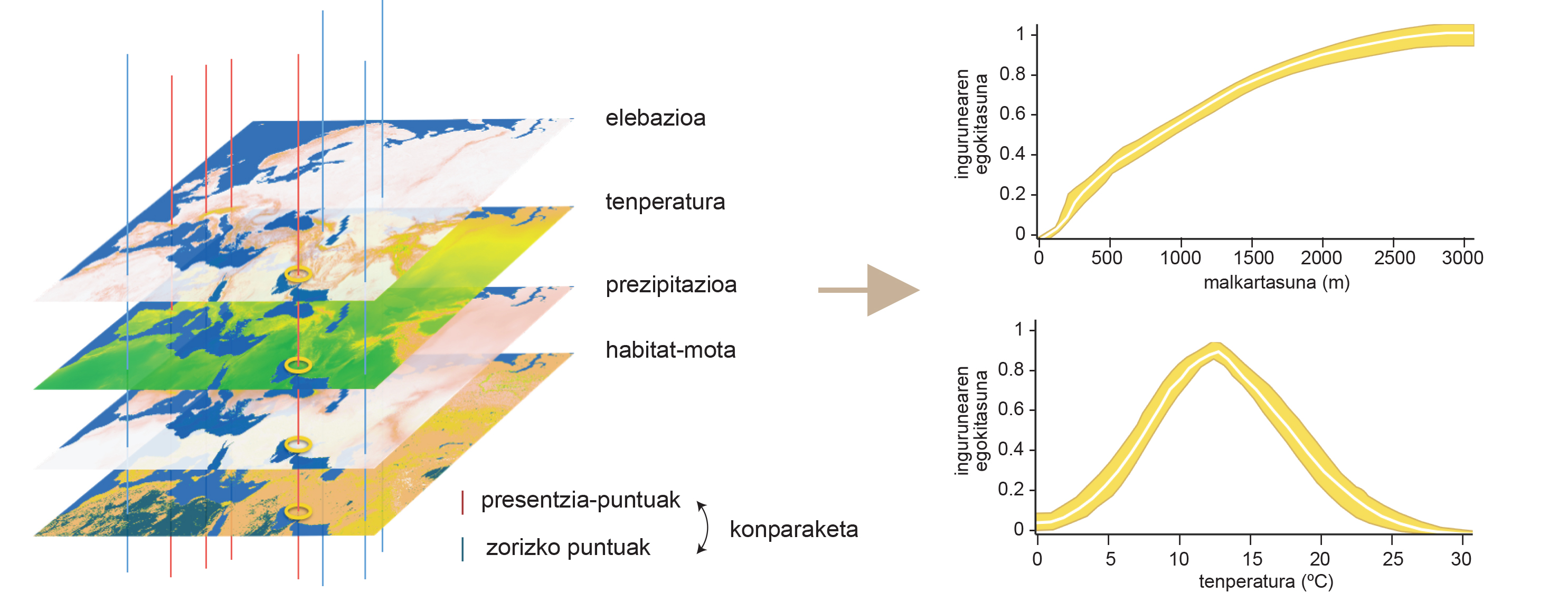
Pour commencer, définissons la niche écologique. Concept XX. Fondée au début du XXe siècle avec deux chercheurs. D'une part, Joseph Grinnell a défini la niche écologique comme un ensemble de conditions environnementales nécessaires à la survie d'une espèce en 1917. Selon Grinnell, dans chaque milieu naturel il y a certaines conditions, et chaque être abrite son espace dans cet ensemble de conditions. Les caractéristiques qui définissent la niche grinnellaise d'un être, que ce soit la température, l'humidité ou les heures de soleil, ne peuvent pas être modifiées par l'être lui-même, c'est-à-dire sont schénopoétiques. Il est donc possible de prédire la distribution spatiale de ces conditions et de les représenter sur des cartes. A partir de ces cartes, il est possible de connaître la niche grinnellaise des espèces par une approche connue sous le nom de modélisation de la niche environnementale (Figure 1). Cette technique compare les conditions environnementales des zones spécifiques dans lesquelles vit une espèce avec celles disponibles dans toute la zone pour déterminer quelles conditions à l'accès de l'animal. Ainsi, par exemple, nous avons appris que la grande oreille alpin vit dans des zones à une température moyenne de 5 à 25 °C par an et qu'elle a besoin topographiquement d'environnements très accidentés pour vivre. Autrement dit, l'herbacé alpin est une espèce qui vit exclusivement dans des zones en pente élevée dans la plage de températures précitée. Cette technique informatique sert également à connaître l'effet relatif de chacune des conditions analysées dans la détermination de la distribution de l'animal. Dans le cas de l'herbacé alpin, nous concluons que l'influence de la topographie est plus significative que celle du climat, c'est-à-dire que la répartition géographique de cette espèce est plus limitée par l'accidentalité du milieu que par la température. Pourquoi ?

Niche écologique rurale
La réponse est au-delà du domaine de la niche grinnellaise, défini par Elton dix ans plus tard. Charles Elton a abordé en 1927 le concept de niche écologique sous un autre angle et a élargi sa signification. Elton s'est concentré sur les ressources environnementales face aux conditions environnementales. En fait, tous les organismes ont besoin de certaines ressources, comme les aliments, le matériel de nidification ou les lieux de placement du réservoir, qui déterminent la niche écologique des organismes. Par conséquent, l'herbacé alpin a besoin de conditions topographiques spécifiques (niche grinnellois), car ce type d'environnements lui offre des ressources spécifiques (niche éltonaise). Contrairement au Grinnelliense, les ressources qui définissent la niche eltonaise interagissent avec l'être lui-même, c'est-à-dire à la différence de la température ou de l'humidité, l'abondance de nourriture peut être modifiée ou épuisée par l'effet de l'être lui-même. Par conséquent, la représentation de ces ressources sur des cartes est pratiquement impossible et nécessite d'autres méthodes pour les connaître.

Pour les chauves-souris, deux sont les principales ressources limitantes : la nourriture et les gardiens. En ce qui concerne les aliments, la grande oreille alpin se nourrit des flancs des zones ouvertes des hautes montagnes, c'est-à-dire des papillons nocturnes qui habitent au-dessus de la côte supérieure des forêts. C'est ce qui a été déduit par des techniques moléculaires basées sur le code-barres ADN (Figure 2). Le code-barres ADN est une partie du génome d'animaux normalisés, une partie du gène appelé COX1, séquencé dans des millions d'animaux et mis à la disposition de tous les chercheurs dans des bases de données géantes. Ainsi, comme si c'était un code à barres d'un livre, en analysant l'ADN des restes d'un animal, comme des poils ou des plumes, nous pouvons savoir à quelle espèce il appartient. Dans le cas des herbivores alpins, nous avons étudié l'ADN de leurs excréments, et nous avons appris quelles espèces d'insectes ont mangé les chiroptères. À partir de cette information, il a été conclu, pour la première fois chez les chauves-souris, que les principaux habitats cinégétiques de l'herbacé alpin sont les pâturages et les chênes alpins.
La prochaine étape pour approfondir la niche eltonaise a été de déterminer l'utilisation qu'ils font des gardes. Pour ce faire, nous capturons les chauves-souris sur les prairies et les chênes de haute montagne et leur implantons des émetteurs radio (Figure 3). Ces appareils émettent des signaux radio chaque seconde et à travers des récepteurs on peut localiser la chauve-souris à tout moment. Pendant les sept jours, plus de quarante chauves-souris ont été suivies et on a déterminé où elles ont été gardées chaque jour. À notre grande surprise, les chauves-souris se sont retrouvées sur de grands murs rocheux situés entre 1500 et 2500 mètres et entre des pierres de gravier, rencontrant un comportement jamais décrit chez les chauves-souris. Après tout ce travail, nous savons donc que la chauve-souris alpine herbacée ne mange pas de sits, mais seulement des espaces ouverts des hautes montagnes, et qu'elle n'est pas conservée partout, mais dans de grands murs de pierre et de gravier. Ces ressources, ainsi que d'autres inconnus, forment la niche eltonaise de la grande oreille alpin.
De niche écologique à histoire évolutive
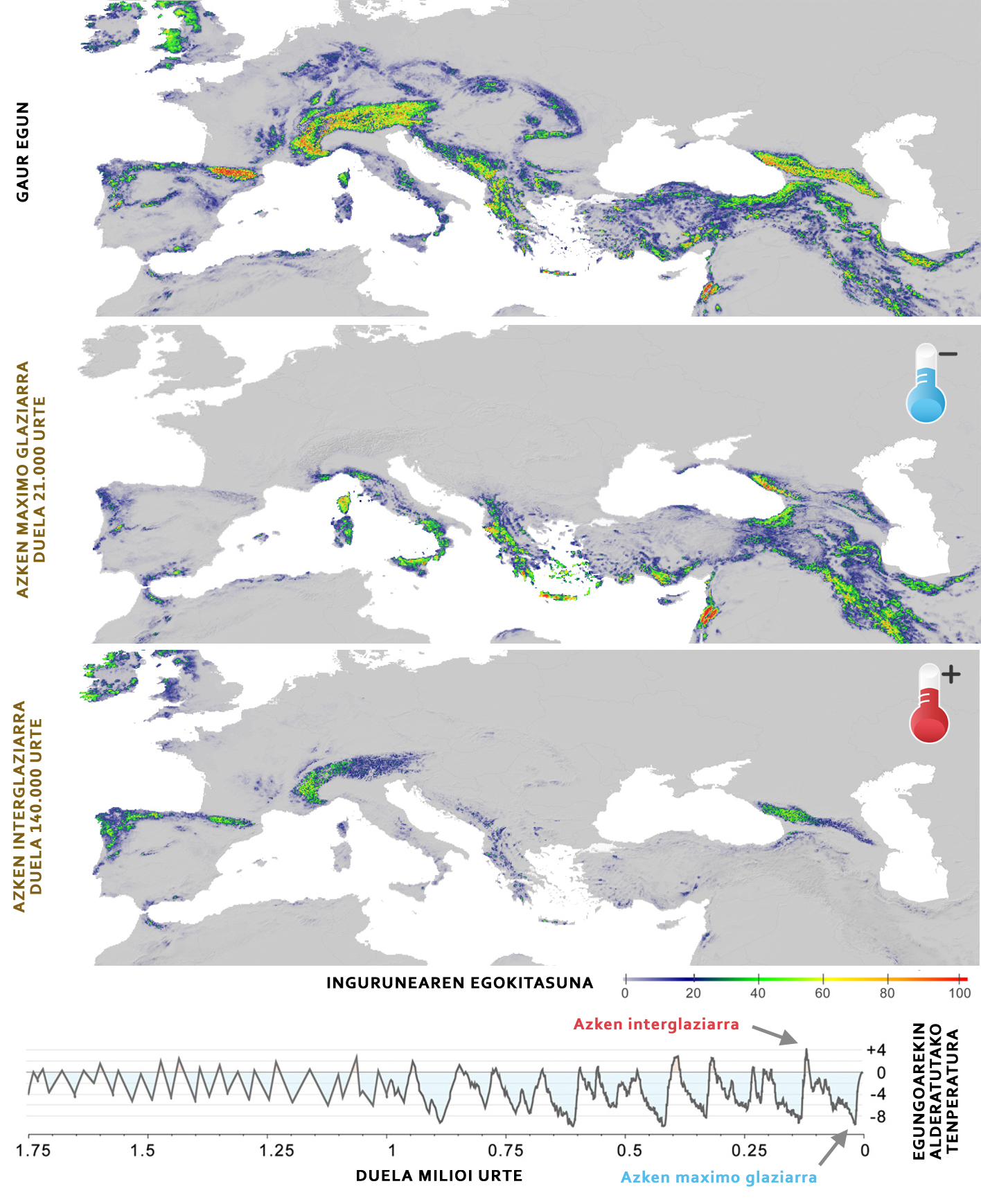
Un des avantages de connaître la niche écologique d'une espèce est qu'elle permet de prédire sa distribution spatiale. C'est-à-dire que, connaissant les conditions requises par une espèce et sa distribution spatiale, nous pouvons prédire quels sont les endroits appropriés pour une espèce et les représenter sur une carte d'adéquation (Figure 4). Si nous savons comment ces conditions et ressources ont évolué tout au long de l'histoire, nous pouvons également évaluer comment les zones appropriées pour l'espèce ont été modifiées dans le temps et, en combinant cette information avec des données moléculaires, reconstruire l'histoire évolutive d'une espèce.
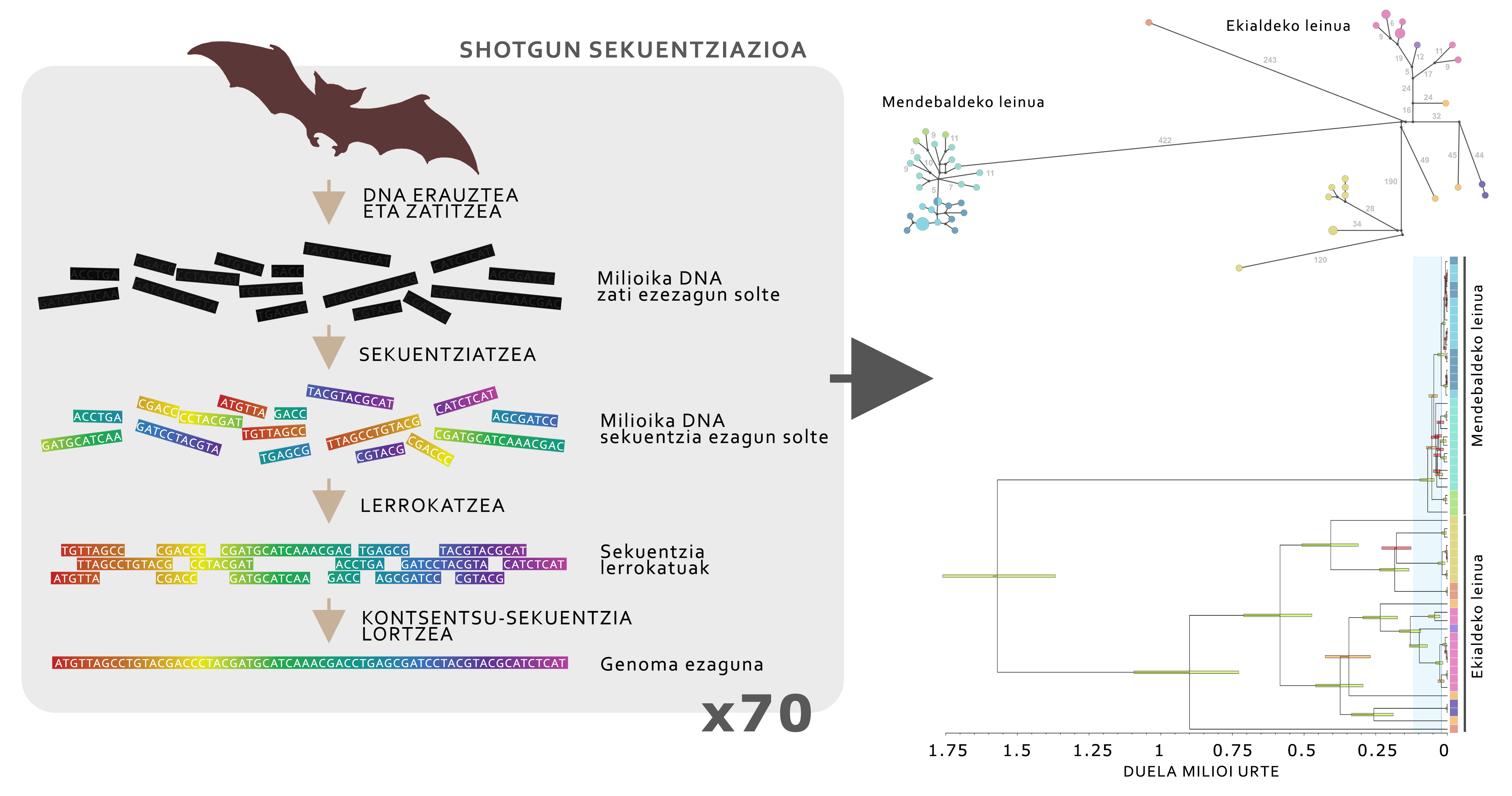
Pour connaître l'histoire évolutive de la grande oreille alpin, nous avons soulevé une série d'hypothèses sur l'évolution de ses populations au fil du temps, sur la base de cartes d'adéquation. Pour tester ces hypothèses, les analyses génomiques de l'ADN ont été utilisées. Après avoir obtenu les échantillons de la membrane sud de 70 animaux répartis dans toute la distribution de l'espèce, on a procédé à l'extraction de l'ADN et au séquençage du génome mitochondrial complet, en utilisant la technique avancée connue comme séquençage de shotgune pour l'étude de l'ADN (Figure 5).
Les données génomiques ont révélé que la grande oreille alpin est divisée en deux lignées principales: Animaux des Pyrénées et des Alpes (lignée occidentale) et toutes les chaînes de montagnes les plus orientales (lignée orientale). Ces deux lignées ont été distribuées il y a environ 1,5 millions d'années et, en raison des glaciations qui ont eu lieu au cours de cette période, les deux populations ont évolué différemment. La lignée occidentale a formé une petite population avec une faible diversité génétique, qui a été fortement touchée par les glaciations, car en refroidissant les températures, l'environnement pyrénéen et alpin devient inapproprié pour la grande oreille alpin (figure 4), parce que ces zones deviennent trop froides. La situation de la lignée orientale, cependant, est complètement différente, car au cours de l'histoire, plusieurs sous-populations sont restées. Dans ce cas, en vivant dans des milieux plus tempérés, lorsque les températures se refroidissent, le degré d'adéquation augmente, ce qui facilite la survie des populations. Cette étude a permis de démontrer que les populations d'une même espèce peuvent avoir des réponses très différentes aux glaciations.
Connaître le passé et le présent pour garantir l’avenir
Il y a quelques années, on connaissait peu de choses sur la chauve-souris oreillette alpin. Nous ne savions pas quelle était sa distribution, ce qu'elle mangeait, où elle était conservée et sans connaître cette information de base il est impossible de prendre les mesures adéquates de conservation. Le travail accompli, cependant, rend la situation actuelle complètement différente. Comme ce fut le cas avec cette espèce, avec l'aide de la technologie développée ces dernières années, nous comprenons le fonctionnement de nombreux éléments et processus de la nature; par la recherche, nous transformons ce qui était autrefois un mystère en une connaissance enracinée par les preuves. Il est maintenant de la responsabilité de tous d'apporter toutes ces informations à un niveau pratique et d'adopter les mesures appropriées pour assurer la survie de la grande oreille alpin et des autres éléments et processus de la nature. Aujourd'hui, comme jamais, il est entre nos mains.
Bibliographie Bibliographie
HEBERT, P.D. (1); CYWINSKA, A.; BALL, S.L. ; DEWAARD, J.R. (2003) : Biological identifications through DNA barcodes. Proceedings of the Royal Society B: Biological Sciences, 270, 313-321.
PHILIPS, S.; DUDIK, M.(2008): Modélisation des distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 31, 161-175.
SPITZENBERGER, F.; STRELKOV, P.; HARING, E. (2003): "Morphology and mitochondrial DNA sequences show that Plecotus alpinus Kiefer Veith, 2002 and Plecotus microdontus Spitzenberger, 2002 are synonyms of Plecotus macrobullaris Kuzjakin, 1965". Nature croate, 12, 39-53.
STEWART, J.R. LISTER, A.M. ; BARNES, I.; DALEN, L. (2010): "Refuges revisited: individualistic responses of species in space and time". Proceedings of the Royal Society B: Biological Sciences, 277, 661-671.
Zu idazle
Zientzia aldizkaria

