

Senar hegoa, allí la cama

Aquest ha estat l'objectiu d'aquesta tesi doctoral que li comptarem: determinar el nínxol ecològic d'un animal desconegut que fins fa poc ha estat fora de la petjada dels científics. Em refereixo a la gran ratapinyada alpina (Plecotus macrobullaris), un animal molt curiós i interessant, com veuràs. Va ser descrit per primera vegada en 2003 als Alps austríacs i durant quatre anys ha estat responsable de les meves suors, maldecaps i gaudi.
En aquell 2010, que ja es veu tan llunyà, em vaig preguntar per què aquesta ratapinyada, al contrari que la resta, està relacionat amb les muntanyes altes. A la recerca d'una resposta, vaig començar a analitzar el nínxol ecològic de l'herbaci alpí al costat dels membres del grup de treball Ecologia i Evolució del Comportament de la UPV/EHU: on es guarda aquest animal? On caça? Quines temperatures pot suportar? Què menja? Cadascuna d'aquestes preguntes fa referència a un eix del nínxol ecològic, i el que comptaré a continuació és l'après en els quatre anys que hem passat buscant respostes.
Nínxol ecològic grinnellés
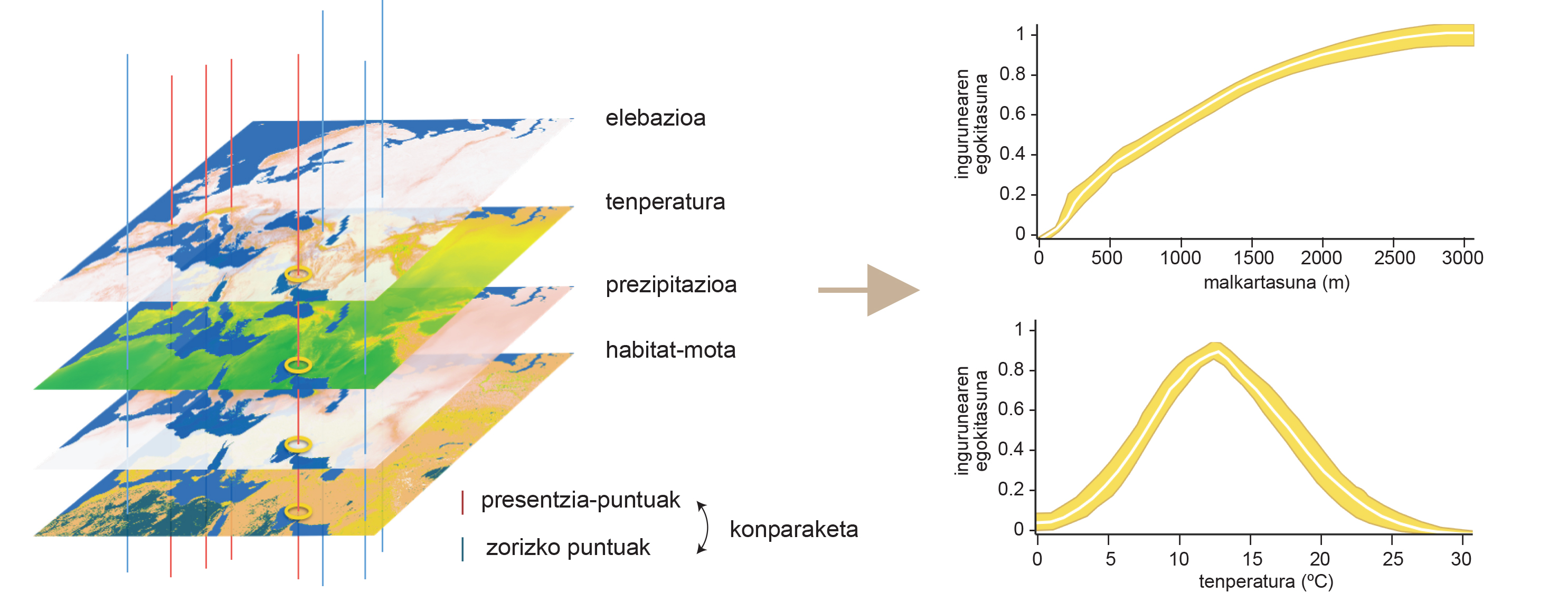
Per a començar definim el nínxol ecològic. Concepte XX. Fundada a principis del segle XX amb dos investigadors. D'una banda, Joseph Grinnell va definir el nínxol ecològic com un conjunt de condicions ambientals necessàries per a la supervivència d'una espècie en 1917. Segons Grinnell, en cada medi natural existeixen unes condicions determinades, i cada ser alberga el seu espai dins d'aquest conjunt de condicions. Les característiques que defineixen el nínxol grinnellés d'un ésser, ja sigui temperatura, humitat o hores de sol, no poden ser modificades pel propi ser, és a dir, són esquenopoéticas. Per això, és possible predir la distribució espacial d'aquestes condicions i representar-les en mapes. A partir d'aquests mapes és possible conèixer el nínxol grinnellés de les espècies mitjançant una aproximació coneguda com a modelització del nínxol ambiental (Figura 1). Aquesta tècnica compara les condicions ambientals de les zones concretes en les quals viu una espècie amb les disponibles en tot l'àrea per a determinar quins de les condicions a les quals s'accedeix l'animal. Així, per exemple, vam conèixer que la gran oïda alpina viu en zones amb una temperatura mitjana d'entre 5 i 25 °C a l'any i que topogràficament necessita d'entorns molt accidentats per a viure. És a dir, l'herbaci alpí és una espècie que viu exclusivament en zones de gran pendent dins del citat rang de temperatures. Aquesta tècnica informàtica serveix també per a conèixer l'efecte relatiu de cadascuna de les condicions analitzades en la determinació de la distribució de l'animal. En el cas de l'herbaci alpí, concloem que la influència de la topografia és més significativa que la del clima, és a dir, que la distribució geogràfica d'aquesta espècie es troba més limitada per l'accidentalitat del mitjà que per la temperatura. Per què?

Nínxol ecològic rural
La resposta està més enllà de l'àmbit del nínxol grinnellés, definit per Elton deu anys després. Charles Elton va abordar en 1927 el concepte de nínxol ecològic des d'una altra perspectiva i va ampliar el seu significat. Elton es va centrar en els recursos ambientals enfront de les condicions ambientals. De fet, tots els organismes necessiten de certs recursos, com els aliments, el material de nidificació o els llocs de col·locació del dipòsit, que determinen el nínxol ecològic dels organismes. Per tant, l'herbaci alpí necessita unes condicions topogràfiques específiques (nínxol grinnellés), ja que aquest tipus d'entorns li ofereix uns recursos concrets (nínxol eltonés). A diferència del Grinnelliense, els recursos que defineixen el nínxol eltonés interactuen amb el propi ser, és a dir, a diferència de la temperatura o de la humitat, l'abundància d'aliments pot alterar-se o esgotar-se per efecte del propi ésser. En conseqüència, la representació d'aquests recursos en mapes és pràcticament impossible i requereix d'altres mètodes per a conèixer-los.

Per a les ratapinyades, dues són els principals recursos limitants: els aliments i les guardes. Pel que fa als aliments, la gran oïda alpina s'alimenta de les faldilles de les zones obertes de les muntanyes altes, és a dir, de les papallones nocturnes que habiten per sobre de la cota superior dels boscos. Això és el que es va deduir mitjançant tècniques moleculars basades en el codi de barres ADN (Figura 2). El codi de barres DNA és una part del genoma d'animals estandarditzats, una part del gen anomenat COX1, seqüenciat en milions d'animals i lloc a la disposició de tots els investigadors en bases de dades gegants. Així, com si fos un codi de barres d'un llibre, analitzant l'ADN de les restes d'un animal, com a pèls o plomes, podem saber a quina espècie pertany. En el cas dels herbívors alpins, estudiem l'ADN dels seus excrements, i aconseguim conèixer quines espècies d'insectes van menjar les ratapinyades. A partir d'aquesta informació, es va concloure, per primera vegada entre les ratapinyades, que els principals hàbitats cinegètics de l'herbaci alpí són les pastures i les rouredes alpines.
El següent pas per a aprofundir en el nínxol eltonés va ser determinar l'ús que fan de les guardes. Per a això, capturem a les ratapinyades en prats i rouredes d'alta muntanya i els implantem emissors de ràdio (figura 3). Aquests aparells emeten senyals de ràdio cada segon i a través d'uns receptors es pot localitzar la ratapinyada a cada moment. Durant els set dies es van seguir més de quaranta ratapinyades i es va determinar on es van guardar cada dia. Per a la nostra sorpresa, les ratapinyades es van trobar en grans murs rocosos situats entre 1.500 i 2.500 metres i entre pedres de graves, trobant un comportament mai descrit entre ratapinyades. Després de tot aquest treball sabem, per tant, que la ratapinyada herbàcia alpina no menja qualsevol sits, sinó només dels espais oberts de les altes muntanyes, i que no es guarda en qualsevol lloc, sinó en grans murs de pedra i graves. Aquests recursos, juntament amb altres desconeguts, formen el nínxol eltonés de la gran oïda alpina.
De nínxol ecològic a història evolutiva
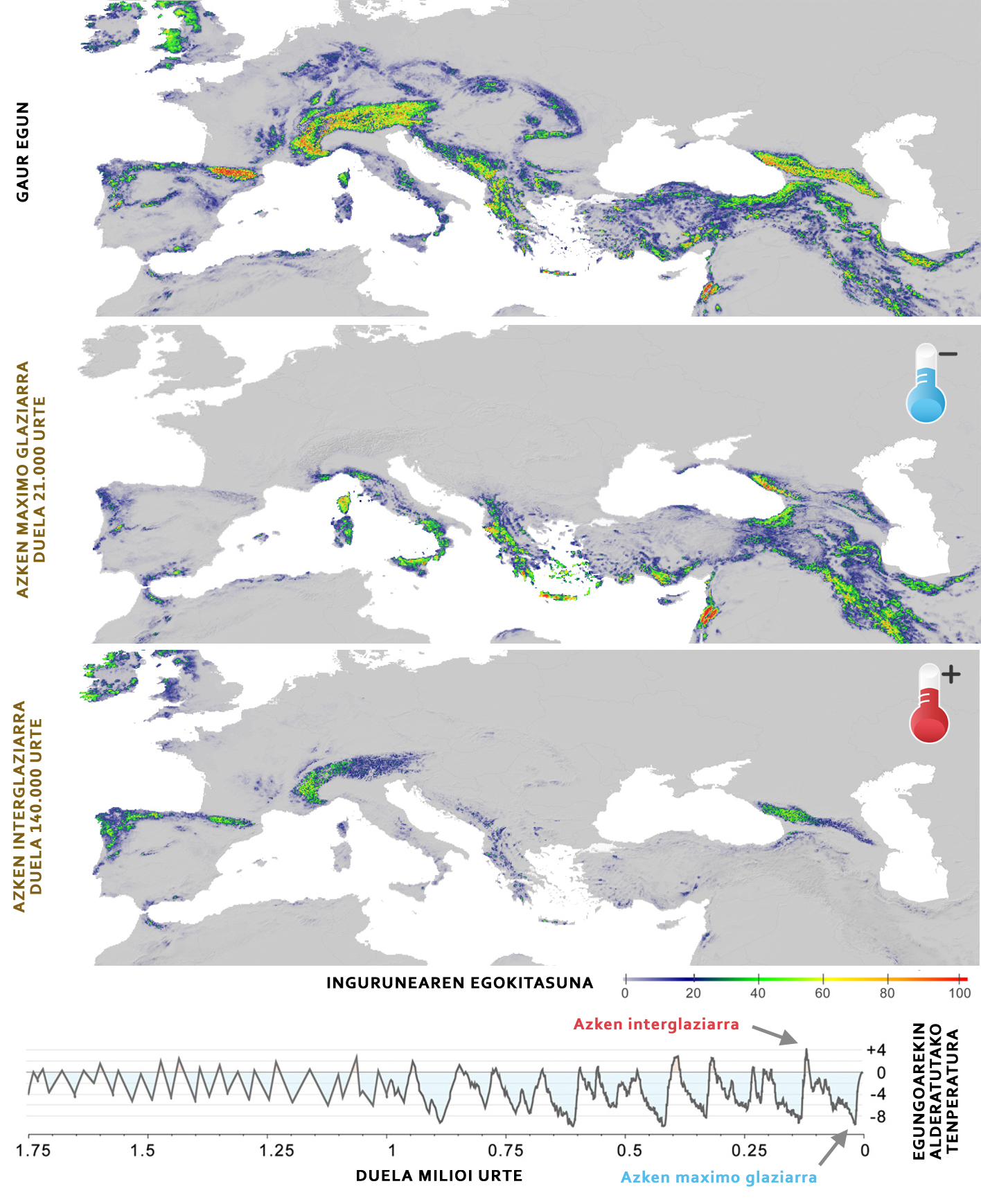
Una dels avantatges de conèixer el nínxol ecològic d'una espècie és que permet predir la seva distribució espacial. És a dir, coneixent les condicions que requereix una espècie i la seva distribució espacial, podem predir quins són els llocs adequats per a una espècie i plasmar-los en un mapa d'idoneïtat (Figura 4). Si sabem com aquestes condicions i recursos han evolucionat al llarg de la història, a més, podem valorar com s'han modificat en el temps les zones apropiades per a l'espècie i, combinant aquesta informació amb dades moleculars, reconstruir la història evolutiva d'una espècie.
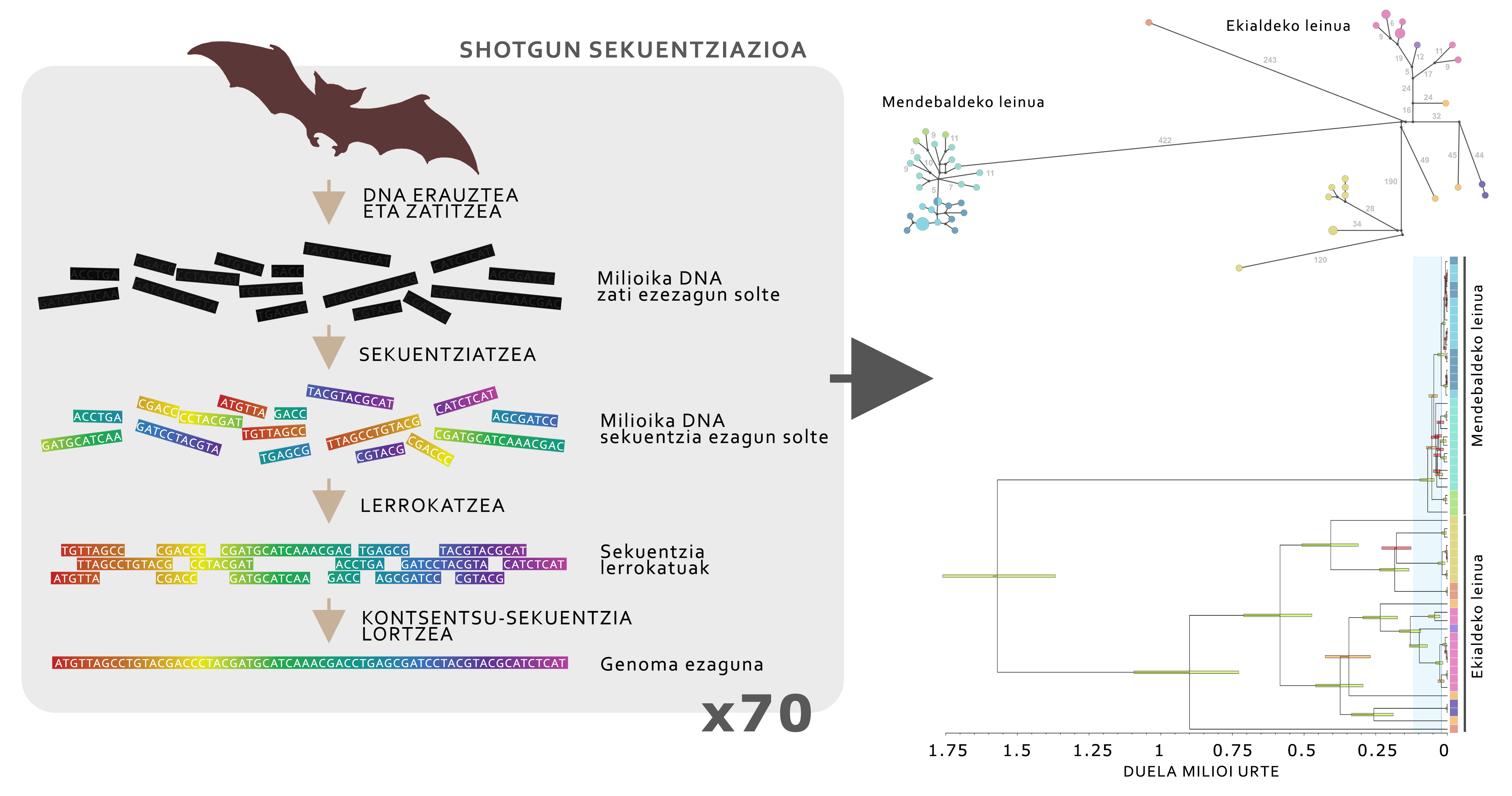
Per a conèixer la història evolutiva de la gran oïda alpina, plantegem una sèrie d'hipòtesi sobre l'evolució de les seves poblacions al llarg del temps, basant-se en mapes d'idoneïtat. Per a testar aquestes hipòtesis es van utilitzar les anàlisis genómicos de l'ADN. Una vegada obtingudes les mostres de la membrana sud de 70 animals repartits en tota la distribució de l'espècie, es va procedir a l'extracció de l'ADN i seqüenciació del genoma mitocondrial complet, utilitzant la tècnica avançada coneguda com a seqüenciació de shotgune per a l'estudi de l'ADN (Figura 5).
Les dades genómicos van revelar que la gran oïda alpina està dividit en dos llinatges principals: Animals dels Pirineus i Alps (llinatge occidental) i tots els de les cadenes muntanyenques més orientals (llinatge oriental). Aquests dos llinatges es van distribuir fa uns 1,5 milions d'anys i, a causa de les glaciacions que s'han produït en aquest període, totes dues poblacions han evolucionat de forma molt diferent. El llinatge occidental ha format una petita població amb una reduïda diversitat genètica, que s'ha vist fortament afectat per les glaciacions, ja que en refredar-se les temperatures, l'entorn pirinenc i alpí es torna inapropiat per a la gran oïda alpina (Figura 4), pel fet que aquestes zones es tornen massa fredes. La situació del llinatge oriental, no obstant això, és completament diferent, ja que al llarg de la història s'han mantingut diverses subpoblaciones. En aquest cas, en viure en mitjans més temperats, quan les temperatures es refreden, augmenta el grau d'idoneïtat, la qual cosa facilita la supervivència de les poblacions. Aquest estudi va servir per a demostrar que les poblacions d'una mateixa espècie poden tenir respostes molt diferents a les glaciacions.
Conèixer el passat i el present per a garantir el futur
Fa uns anys es coneixia poc sobre la ratapinyada orelluda alpina. No sabíem quina era la seva distribució, què menjava, on es guardava i sense conèixer aquesta informació bàsica és impossible prendre les mesures adequades de conservació. El treball realitzat, no obstant això, fa que la situació actual sigui completament diferent. Igual que va ocórrer amb aquesta espècie, amb l'ajuda de la tecnologia desenvolupada en els últims anys, estem comprenent el funcionament de molts elements i processos de la naturalesa; mitjançant la recerca estem convertint el que abans era un misteri en un coneixement arrelat per les evidències. Ara és responsabilitat de tots portar tota aquesta informació a un nivell pràctic i adoptar les mesures adequades per a garantir la supervivència de la gran oïda alpina i dels altres elements i processos de la naturalesa. Avui, com mai, és a les nostres mans.
Bibliografia
HEBERT, P.D. (1); CYWINSKA, A.; BALL, S.L. ; DEWAARD, J.R. (2003): Biological identifications through DNA barcodes. Proceedings of the Royal Society B: Biological Sciences, 270, 313-321.
PHILIPS, S.; DUDIK, M.(2008): Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 31, 161-175.
SPITZENBERGER, F.; STRELKOV, P.; HARING, E. (2003): "Morphology and mitochondrial DNA sequences xou that Plecotus alpinus Kiefer & Veith, 2002 and Plecotus microdontus Spitzenberger, 2002 llauri synonyms of Plecotus macrobullaris Kuzjakin, 1965". Naturalesa Croática, 12, 39-53.
STEWART, J.R. LISTER, A. m.; BARNES, I.; DALEN, L. (2010): "Refugia revisited: individualistic responses of species in space and estafi". Proceedings of the Royal Society B: Biological Sciences, 277, 661-671.
Zu idazle
Zientzia aldizkaria

