

Pola noite son capaces de velo en cores, pero como?

A visión é un dos factores máis importantes na supervivencia dun individuo, imprescindible na persecución de presas, na evitación de depredadores e no proceso de descubrimento da súa parella. Presenta una gran variabilidade e a súa capacidade de adaptación a diferentes situacións, así como á súa evolución. Así, algunhas estruturas do ollo, como o tapetum luciduma, chegaron a ser funcionalmente iguais en liñaxes sen estreitas relacións evolutivas.
As estruturas imprescindibles paira poder velo son os fotoreceptores situados na retina do ollo. Poden ser conos (visión de cores) e bastóns (intensidade da luz). Nos conos aparecerá a molécula de opsina, que pode ser de tres tipos: As S opsinas son sensibles á luz de lonxitude de onda curta, as M opsinas á lonxitude de onda intermedia e as L opsinas ás grandes lonxitudes de onda, permitindo visualizar a cor azul-violeta, verde e vermello respectivamente. Os tipos de opsina que terá cada ser estarán xeneticamente determinados. Nos paus, en lugar de opsina, aparece a rodopsina, que dependendo da intensidade lumínica da zona terá distinto nivel de actividade. A pesar de estar inactivo en iluminacións moi altas, é moi útil paira aproveitar a escasa iluminación existente.
A selección natural, como mecanismo evolutivo, actúa de maneira diferente en cada momento e lugar concreto e só avanzan os xenes que favorece ao longo do tempo. En orixe, a visión na liñaxe dos mamíferos era dicromata (distinguían dúas cores), pero variou en función da evolución de cada liña evolutiva, segundo as necesidades de cada un e a influencia da selección. Na situación orixinal, a opsina S sempre aparecía e con ela a opsina M ou L. Nos cetáceos actuais, por exemplo, o xene que codifica a opsina S desapareceu e son monocromatas (veno nunha única gama de cores). Na liñaxe humana, pola súa banda, nos xenes que determinaban a dicromata orixinal produciuse una duplicación e o xene duplicado habería desenvolvido una nova función, dando lugar á situación actual a unha tricromata (o tres opsinas están á vez na retina, sendo capaces de distinguir tres gamas de cores).
Dentro da variabilidade visual, hai situacións características como o modelo de visión desenvolvido en animais nocturnos. Paira estes animais é moi importante que a pouca luz nocturna (estrelas, lúa...) diríxase á retina e paira iso os paus da retina sexan o máis sensibles posible. Así, Solovei et ao. descubriron en 2009 que, nos paus de animais nocturnos, o patrón de distribución do ADN que aparece nos bastóns do resto de animais está investido, o que permite una maior produción de rodopsina, aumentando a sensibilidade á luz.

Ademais de desenvolver a visión nocturna, existen tamén animais que desenvolveron una situación máis extrema: os capaces de velo de noite e de cores. Sempre se considerou que os cambios visuais producidos durante a evolución dos mamíferos están relacionados coa persecución de alimentos e presas. Así, relacionouse a dicromacia cos animais do día, mentres que nos animais nocturnos considerouse que a cor era pouco importante. Curiosamente, con todo, as investigacións actuais non apoian esta idea, xa que se ha visto que algúns mamíferos nocturnos poden distinguir as cores. Aínda que non alcanzan a resolución das actuais tricromatas, manteñen dúas opsinas.
En relación ás razóns polas que se mantivo a existencia de dúas opsinas, os investigadores formularon dúas hipótese. Segundo a primeira, nestes animais a dicromacia non é tan vantaxosa en si mesma, senón a característica plesiomorfológica (herdada dos antepasados) dos animais recentemente convertidos en nocturnos. Na segunda hipótese, a dicromacia sería una característica vantaxosa paira os animais nocturnos, ou os xenes de opínaa, ademais dunha función relacionada coa cor, cumprirían algunha outra. Os estudos demostran que as opsinas M e L mantivéronse sen grandes cambios nalgunhas liñaxes nocturnas respecto do seu estado orixinal, como o aye-aye ( Daubentonia madagascariensis ) e algúns morcegos.
Aye-ayea
En 2007, Perry et ao. compararon a secuencia de ADN do aye-aye coa doutros lemuriformes paira observar si houbo ou non cambio nos xenes da opsina e, a partir de aí, dar resposta á capacidade de visión nocturna e a cor. Así, nas opsinas do aye-ayea non se detectaron cambios respecto da dicromática orixinal, e pode pensarse que a separación de cores suporía una vantaxe paira estes animais nocturnos e que os cambios producidos, ao non ser favorados, desaparecían.

O aye-ayea, que se repartiu desde hai moito tempo entre o resto do grupo, desenvolveu durante todo este tempo una serie de características paira adaptarse ao seu nicho ecolóxico. Con todo, os xenes opsina mantiveron o seu estado orixinal ao longo do tempo, reforzando novamente a hipótese sobre a súa funcionalidade.
Morcegos
Os morcegos divídense en dous grandes grupos: As megachiropteras frutajales ( Yinpterochiroptera ) e as insectívoras Microchiropteras ( Yangochiroptera ) desenvolveron diferentes formas de ecolocalización, xa que as súas necesidades son diferentes. As primeiras teñen una boa evolución visual e olfativa e o eco-localización desenvolvida é de tipo low-duty-cycle. Os sons que emiten son de curta duración e non se solapan con eco. No caso dos segundos, a ecolocalización é laríngea e as ondas emítense constantemente, superponiéndolas coas ondas que están a retornar.
Coa diferente eficiencia de ambas as formas de ecolocalización, e cos ollos menos desenvolvidos en Microchiropter, adóitase pensar que estes morcegos se basean no eco-localización e que non teñen moi boa visión. Con todo, a opsina S mantívose en ambos os grupos, sendo capaces de ver a luz ultravioleta e, por tanto, non son cegas, como se adoita dicir a miúdo. Una das razóns polas que os morcegos poden ver a luz ultravioleta pode estar relacionada coas ás dos lepidópteros gautanos que forman parte da súa dieta, dos cales o 80% reflicten a luz ultravioleta. Doutra banda, outra proba da utilidade da vista está na forma de caza dos morcegos. De feito, tras achegarse á presa a través das ondas de ecolocalización, no último paso utilizan a vista, xa que ao ser capaces de detectar as ondas os insectos evitaríanse.
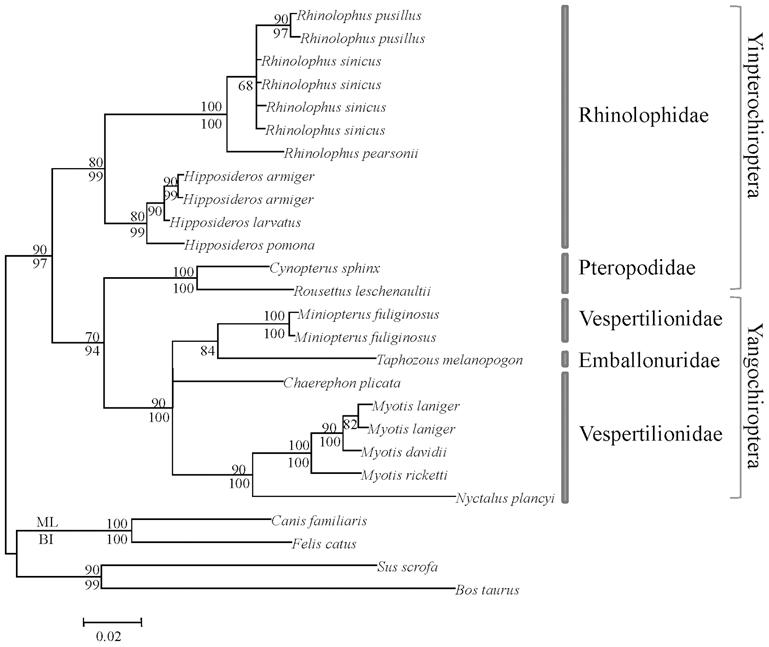
En cambio, en Megachiropter, debido principalmente á súa dieta, o eco-localización é menos importante. E é que paira detectar cando as flores están cheas de néctares e os froitos maduros, o olfacto e a vista son fundamentais.
Segundo os resultados obtidos nos estudos xenéticos, en todos os grupos de morcegos mantívose a opsina S, así como a opsina M ou L, destacando a segunda. A elección polo mantemento do alelo L pode estar relacionada coa visión da cor vermella, indicador da madurez de moitos froitos. As árbores filogenéticos que se constrúen cos datos obtidos nos estudos realizados con estes xenes, ademais, reflicten adecuadamente a evolución dada na liñaxe dos morcegos e confirman a actual agrupación.
Segundo o visto, a visión tivo diferentes formas de desenvolvemento, chegando a situacións a miúdo converxentes en función das necesidades e comportamento etoecológico dos animais. Con todo, nalgúns animais nocturnos, non mamíferos e filogenéticamente afastados dos mesmos, tamén se atopou a capacidade de diferenciar as cores pola noite; o gecko e algúns sits da familia Sphingidae son capaces de distinguir 6 cores, como a luz da lúa ou as estrelas. Aínda queda moito por investigar neste campo.
Referencias
Zu idazle
Zientzia aldizkaria

