

A la nit són capaços de veure-ho en colors, però com?

La visió és un dels factors més importants en la supervivència d'un individu, imprescindible en la persecució de preses, en l'evitació de depredadors i en el procés de descobriment de la seva parella. Presenta una gran variabilitat i la seva capacitat d'adaptació a diferents situacions, així com a la seva evolució. Així, algunes estructures de l'ull, com el tapetum luciduma, han arribat a ser funcionalment iguals en llinatges sense estretes relacions evolutives.
Les estructures imprescindibles per a poder veure-ho són els fotoreceptores situats en la retina de l'ull. Poden ser cons (visió de colors) i bastons (intensitat de la llum). En els cons apareixerà la molècula d'opsina, que pot ser de tres tipus: Les S opsinas són sensibles a la llum de longitud d'ona curta, les M opsinas a la longitud d'ona intermèdia i les L opsinas a les grans longituds d'ona, permetent visualitzar el color blau-violeta, verda i vermell respectivament. Els tipus d'opsina que tindrà cada ser estaran genèticament determinats. En els pals, en lloc d'opsina, apareix la rodopsina, que depenent de la intensitat lumínica de la zona tindrà diferent nivell d'activitat. Malgrat estar inactiu en il·luminacions molt altes, és molt útil per a aprofitar l'escassa il·luminació existent.
La selecció natural, com a mecanisme evolutiu, actua de manera diferent a cada moment i lloc concret i només avancen els gens que afavoreix al llarg del temps. En origen, la visió en el llinatge dels mamífers era dicromata (distingien dos colors), però ha variat en funció de l'evolució de cada línia evolutiva, segons les necessitats de cadascun i la influència de la selecció. En la situació original, l'opsina S sempre apareixia i amb ella l'opsina M o L. En els cetacis actuals, per exemple, el gen que codifica l'opsina S ha desaparegut i són monocromatas (ho veuen en una única gamma de colors). En el llinatge humà, per part seva, en els gens que determinaven la dicromata original s'hauria produït una duplicació i el gen duplicat hauria desenvolupat una nova funció, donant lloc a la situació actual a una tricromata (les tres opsinas estan alhora en la retina, sent capaces de distingir tres gammes de colors).
Dins de la variabilitat visual, hi ha situacions característiques com el model de visió desenvolupat en animals nocturns. Per a aquests animals és molt important que la poca llum nocturna (estrelles, lluna...) es dirigeixi a la retina i per a això els pals de la retina siguin el més sensibles possible. Així, Solovei et al. van descobrir en 2009 que, en els pals d'animals nocturns, el patró de distribució de l'ADN que apareix en els bastons de la resta d'animals està invertit, la qual cosa permet una major producció de rodopsina, augmentant la sensibilitat a la llum.

A més de desenvolupar la visió nocturna, existeixen també animals que han desenvolupat una situació més extrema: els capaços de veure-ho de nit i de colors. Sempre s'ha considerat que els canvis visuals produïts durant l'evolució dels mamífers estan relacionats amb la persecució d'aliments i preses. Així, s'ha relacionat la dicromacia amb els animals del dia, mentre que en els animals nocturns s'ha considerat que el color era poc important. Curiosament, no obstant això, les recerques actuals no donen suport a aquesta idea, ja que s'ha vist que alguns mamífers nocturns poden distingir els colors. Encara que no aconsegueixen la resolució de les actuals tricromatas, mantenen dues opsinas.
En relació a les raons per les quals s'ha mantingut l'existència de dues opsinas, els investigadors han formulat dues hipòtesis. Segons la primera, en aquests animals la dicromacia no és tan avantatjosa en si mateixa, sinó la característica plesiomorfológica (heretada dels avantpassats) dels animals recentment convertits en nocturns. En la segona hipòtesi, la dicromacia seria una característica avantatjosa per als animals nocturns, o els gens de l'opina, a més d'una funció relacionada amb el color, complirien alguna altra. Els estudis demostren que les opsinas M i L s'han mantingut sense grans canvis en alguns llinatges nocturns respecte al seu estat original, com l'aye-aye ( Daubentonia madagascariensis ) i algunes ratapinyades.
Aye-ayea
En 2007, Perry et al. van comparar la seqüència d'ADN de l'aye-aye amb la d'uns altres lemuriformes per a observar si va haver-hi o no canvi en els gens de l'opsina i, a partir d'aquí, donar resposta a la capacitat de visió nocturna i a color. Així, en les opsinas de l'aye-ayea no es van detectar canvis respecte a la dicromática original, i pot pensar-se que la separació de colors suposaria un avantatge per a aquests animals nocturns i que els canvis produïts, al no ser favorados, desapareixien.

L'aye-ayea, que es va repartir des de fa molt temps entre la resta del grup, ha desenvolupat durant tot aquest temps una sèrie de característiques per a adaptar-se al seu nínxol ecològic. No obstant això, els gens opsina han mantingut el seu estat original al llarg del temps, reforçant novament la hipòtesi sobre la seva funcionalitat.
Ratapinyades
Les ratapinyades es divideixen en dos grans grups: Les megachiropteras frutajales ( Yinpterochiroptera ) i les insectívores Microchiropteras ( Yangochiroptera ) han desenvolupat diferents formes d'ecolocalización, ja que les seves necessitats són diferents. Les primeres tenen una bona evolució visual i olfactòria i l'eco-localització desenvolupada és de tipus low-duty-cycle. Els sons que emeten són de curta durada i no se solapen amb ressò. En el cas dels segons, l'ecolocalización és laríngia i les ones s'emeten constantment, superposant-les amb les ones que estan retornant.
Amb la diferent eficiència de totes dues formes d'ecolocalización, i amb els ulls menys desenvolupats en Microchiropter, se sol pensar que aquestes ratapinyades es basen en l'eco-localització i que no tenen molt bona visió. No obstant això, l'opsina S s'ha mantingut en tots dos grups, sent capaços de veure la llum ultraviolada i, per tant, no són cegues, com se sol dir sovint. Una de les raons per les quals les ratapinyades poden veure la llum ultraviolada pot estar relacionada amb les ales dels lepidòpters gautanos que formen part de la seva dieta, dels quals el 80% reflecteixen la llum ultraviolada. D'altra banda, una altra prova de la utilitat de la vista està en la forma de caça de les ratapinyades. De fet, després d'acostar-se a la presa a través de les ones d'ecolocalización, en l'últim pas utilitzen la vista, ja que en ser capaces de detectar les ones els insectes s'evitarien.
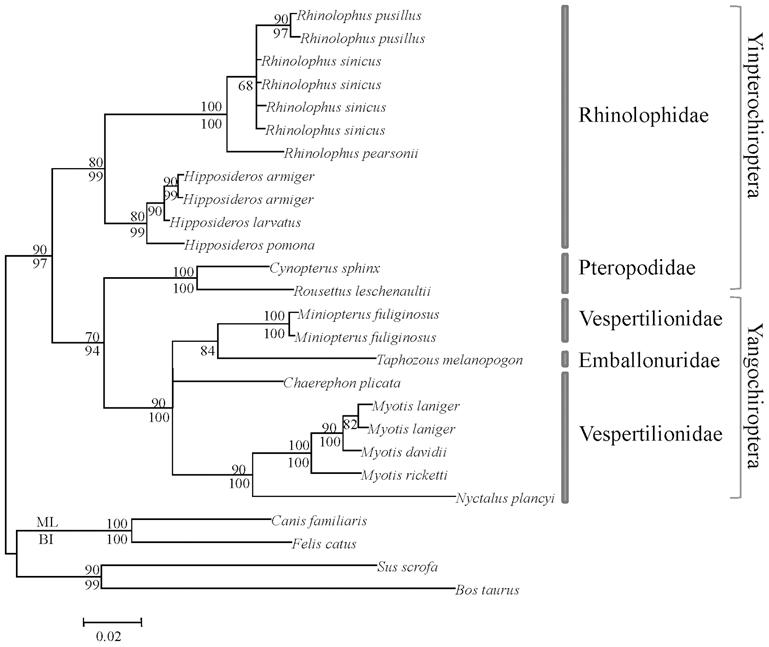
En canvi, en Megachiropter, degut principalment a la seva dieta, l'eco-localització és menys important. I és que per a detectar quan les flors estan plenes de nèctars i els fruits madurs, l'olfacte i la vista són fonamentals.
Segons els resultats obtinguts en els estudis genètics, en tots els grups de ratapinyades s'ha mantingut l'opsina S, així com l'opsina M o L, destacant la segona. L'elecció pel manteniment de l'al·lel L pot estar relacionada amb la visió del color vermell, indicador de la maduresa de molts fruits. Els arbres filogenètics que es construeixen amb les dades obtingudes en els estudis realitzats amb aquests gens, a més, reflecteixen adequadament l'evolució donada en el llinatge de les ratapinyades i confirmen l'actual agrupació.
Segons el vist, la visió ha tingut diferents formes de desenvolupament, arribant a situacions sovint convergents en funció de les necessitats i comportament etoecológico dels animals. No obstant això, en alguns animals nocturns, no mamífers i filogenéticamente allunyats d'aquests, també s'ha trobat la capacitat de diferenciar els colors a la nit; el gecko i alguns sits de la família Sphingidae són capaces de distingir 6 colors, com la llum de la lluna o les estrelles. Encara queda molt per investigar en aquest camp.
Referències
Zu idazle
Zientzia aldizkaria

