

Por la noche son capaces de verlo en colores, pero ¿cómo?

La visión es uno de los factores más importantes en la supervivencia de un individuo, imprescindible en la persecución de presas, en la evitación de depredadores y en el proceso de descubrimiento de su pareja. Presenta una gran variabilidad y su capacidad de adaptación a diferentes situaciones, así como a su evolución. Así, algunas estructuras del ojo, como el tapetum luciduma, han llegado a ser funcionalmente iguales en linajes sin estrechas relaciones evolutivas.
Las estructuras imprescindibles para poder verlo son los fotoreceptores situados en la retina del ojo. Pueden ser conos (visión de colores) y bastones (intensidad de la luz). En los conos aparecerá la molécula de opsina, que puede ser de tres tipos: Las S opsinas son sensibles a la luz de longitud de onda corta, las M opsinas a la longitud de onda intermedia y las L opsinas a las grandes longitudes de onda, permitiendo visualizar el color azul-violeta, verde y rojo respectivamente. Los tipos de opsina que tendrá cada ser estarán genéticamente determinados. En los palos, en lugar de opsina, aparece la rodopsina, que dependiendo de la intensidad lumínica de la zona tendrá distinto nivel de actividad. A pesar de estar inactivo en iluminaciones muy altas, es muy útil para aprovechar la escasa iluminación existente.
La selección natural, como mecanismo evolutivo, actúa de manera diferente en cada momento y lugar concreto y sólo avanzan los genes que favorece a lo largo del tiempo. En origen, la visión en el linaje de los mamíferos era dicromata (distinguían dos colores), pero ha variado en función de la evolución de cada línea evolutiva, según las necesidades de cada uno y la influencia de la selección. En la situación original, la opsina S siempre aparecía y con ella la opsina M o L. En los cetáceos actuales, por ejemplo, el gen que codifica la opsina S ha desaparecido y son monocromatas (lo ven en una única gama de colores). En el linaje humano, por su parte, en los genes que determinaban la dicromata original se habría producido una duplicación y el gen duplicado habría desarrollado una nueva función, dando lugar a la situación actual a una tricromata (las tres opsinas están a la vez en la retina, siendo capaces de distinguir tres gamas de colores).
Dentro de la variabilidad visual, hay situaciones características como el modelo de visión desarrollado en animales nocturnos. Para estos animales es muy importante que la poca luz nocturna (estrellas, luna...) se dirija a la retina y para ello los palos de la retina sean lo más sensibles posible. Así, Solovei et al. descubrieron en 2009 que, en los palos de animales nocturnos, el patrón de distribución del ADN que aparece en los bastones del resto de animales está invertido, lo que permite una mayor producción de rodopsina, aumentando la sensibilidad a la luz.

Además de desarrollar la visión nocturna, existen también animales que han desarrollado una situación más extrema: los capaces de verlo de noche y de colores. Siempre se ha considerado que los cambios visuales producidos durante la evolución de los mamíferos están relacionados con la persecución de alimentos y presas. Así, se ha relacionado la dicromacia con los animales del día, mientras que en los animales nocturnos se ha considerado que el color era poco importante. Curiosamente, sin embargo, las investigaciones actuales no apoyan esta idea, ya que se ha visto que algunos mamíferos nocturnos pueden distinguir los colores. Aunque no alcanzan la resolución de las actuales tricromatas, mantienen dos opsinas.
En relación a las razones por las que se ha mantenido la existencia de dos opsinas, los investigadores han formulado dos hipótesis. Según la primera, en estos animales la dicromacia no es tan ventajosa en sí misma, sino la característica plesiomorfológica (heredada de los antepasados) de los animales recientemente convertidos en nocturnos. En la segunda hipótesis, la dicromacia sería una característica ventajosa para los animales nocturnos, o los genes de la opina, además de una función relacionada con el color, cumplirían alguna otra. Los estudios demuestran que las opsinas M y L se han mantenido sin grandes cambios en algunos linajes nocturnos respecto a su estado original, como el aye-aye ( Daubentonia madagascariensis ) y algunos murciélagos.
Aye-ayea
En 2007, Perry et al. compararon la secuencia de ADN del aye-aye con la de otros lemuriformes para observar si hubo o no cambio en los genes de la opsina y, a partir de ahí, dar respuesta a la capacidad de visión nocturna y a color. Así, en las opsinas del aye-ayea no se detectaron cambios respecto a la dicromática original, y puede pensarse que la separación de colores supondría una ventaja para estos animales nocturnos y que los cambios producidos, al no ser favorados, desaparecían.

El aye-ayea, que se repartió desde hace mucho tiempo entre el resto del grupo, ha desarrollado durante todo este tiempo una serie de características para adaptarse a su nicho ecológico. Sin embargo, los genes opsina han mantenido su estado original a lo largo del tiempo, reforzando nuevamente la hipótesis sobre su funcionalidad.
Murciélagos
Los murciélagos se dividen en dos grandes grupos: Las megachiropteras frutajales ( Yinpterochiroptera ) y las insectívoras Microchiropteras ( Yangochiroptera ) han desarrollado diferentes formas de ecolocalización, ya que sus necesidades son diferentes. Las primeras tienen una buena evolución visual y olfativa y la eco-localización desarrollada es de tipo low-duty-cycle. Los sonidos que emiten son de corta duración y no se solapan con eco. En el caso de los segundos, la ecolocalización es laríngea y las ondas se emiten constantemente, superponiéndolas con las ondas que están retornando.
Con la diferente eficiencia de ambas formas de ecolocalización, y con los ojos menos desarrollados en Microchiropter, se suele pensar que estos murciélagos se basan en la eco-localización y que no tienen muy buena visión. Sin embargo, la opsina S se ha mantenido en ambos grupos, siendo capaces de ver la luz ultravioleta y, por tanto, no son ciegas, como se suele decir a menudo. Una de las razones por las que los murciélagos pueden ver la luz ultravioleta puede estar relacionada con las alas de los lepidópteros gautanos que forman parte de su dieta, de los cuales el 80% reflejan la luz ultravioleta. Por otro lado, otra prueba de la utilidad de la vista está en la forma de caza de los murciélagos. De hecho, tras acercarse a la presa a través de las ondas de ecolocalización, en el último paso utilizan la vista, ya que al ser capaces de detectar las ondas los insectos se evitarían.
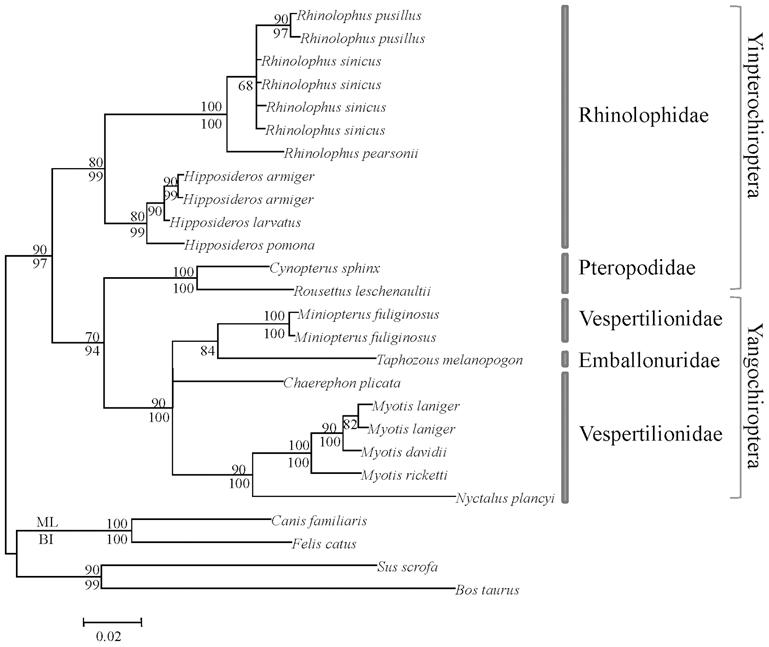
En cambio, en Megachiropter, debido principalmente a su dieta, la eco-localización es menos importante. Y es que para detectar cuándo las flores están llenas de néctares y los frutos maduros, el olfato y la vista son fundamentales.
Según los resultados obtenidos en los estudios genéticos, en todos los grupos de murciélagos se ha mantenido la opsina S, así como la opsina M o L, destacando la segunda. La elección por el mantenimiento del alelo L puede estar relacionada con la visión del color rojo, indicador de la madurez de muchos frutos. Los árboles filogenéticos que se construyen con los datos obtenidos en los estudios realizados con estos genes, además, reflejan adecuadamente la evolución dada en el linaje de los murciélagos y confirman la actual agrupación.
Según lo visto, la visión ha tenido diferentes formas de desarrollo, llegando a situaciones a menudo convergentes en función de las necesidades y comportamiento etoecológico de los animales. Sin embargo, en algunos animales nocturnos, no mamíferos y filogenéticamente alejados de los mismos, también se ha encontrado la capacidad de diferenciar los colores por la noche; el gecko y algunos sits de la familia Sphingidae son capaces de distinguir 6 colores, como la luz de la luna o las estrellas. Todavía queda mucho por investigar en este campo.
Referencias
Zu idazle
Zientzia aldizkaria

