

Los peces hablan de ecología durante las comidas
 400
400
El protagonismo de las anchoas en el Golfo de Bizkaia durante la última década, viene de la mano de los pescadores vascos. Sin embargo, más allá de las anchoas, en general se ha puesto de manifiesto la disminución de la cantidad de peces disponibles en el mar, lo que ha influido directamente en la actividad de los pescadores. La investigación pesquera nos ha demostrado que, por un lado, el cambio climático y, por otro, el exceso de pesca, la situación de la pesca puede empeorar en el futuro si no cuidamos el ecosistema marino. Así, conocer bien el funcionamiento del ecosistema es de vital importancia para garantizar una pesca sostenible, teniendo en cuenta que el conocimiento que tenemos al respecto sigue siendo limitado en muchos aspectos. Por ejemplo, se sabe que las principales especies de peces pelágicos tienen como principal fuente de alimento el zooplancton, y que los cambios en la abundancia y composición del zooplancton derivados de los climas y estructuras oceanográficas pueden afectar directamente al reclutamiento de los peces. Por lo tanto, se puede afirmar que la alteración de la composición y abundancia del alimento de los peces, en la medida en que ello repercuta directamente en la cadena trófica, puede tener consecuencias trágicas en la abundancia y difusión de los mismos.
En este sentido, el objetivo principal de esta tesis es analizar qué comen las principales especies pelágicas menores presentes en el golfo de Bizkaia. Así, además de la determinación de la dieta, se han analizado las posibles limitaciones y la competitividad de los peces en la alimentación, estableciendo superposiciones de dieta en función del espacio y el tiempo que se consideren imprescindibles en la supervivencia de la población de estas especies y, por tanto, en la regulación, y las interacciones entre especies. Es la primera vez que se lleva a cabo una investigación de este tipo en el Golfo de Bizkaia, en la que se han abierto cerca de 800 estómagos de pescado, en los que se han identificado individualmente más de 90.000 presas y medido más de...

Paradoja de la alimentación de la anchoa juvenil
Como se ha mencionado anteriormente, si bien la anchoa europea ha recibido el mayor interés de la investigación científica durante la prohibición de la pesca de esta especie (2005-2010), en este período se trata de la primera investigación que estudia la ecología trófica de la anchoa en el golfo de Bizkaia. De este modo, se ha comprobado que los juveniles de anchoa presentan un comportamiento alimentario particular, sin necesidad de profundizar en las características de su dieta. De hecho, entre 2003 y 2010, en otoño, analizando el peso del estómago de las anchoas jóvenes tomadas para muestras (grado de llenado del estómago según zonas), tanto fuera de la plataforma continental como en el talud continental, al ser la concentración de zooplancton disponible menor, el peso del estómago de las anchoas jóvenes es mayor (medido este peso respecto al peso total del pescado) que en el interior de la plataforma continental.
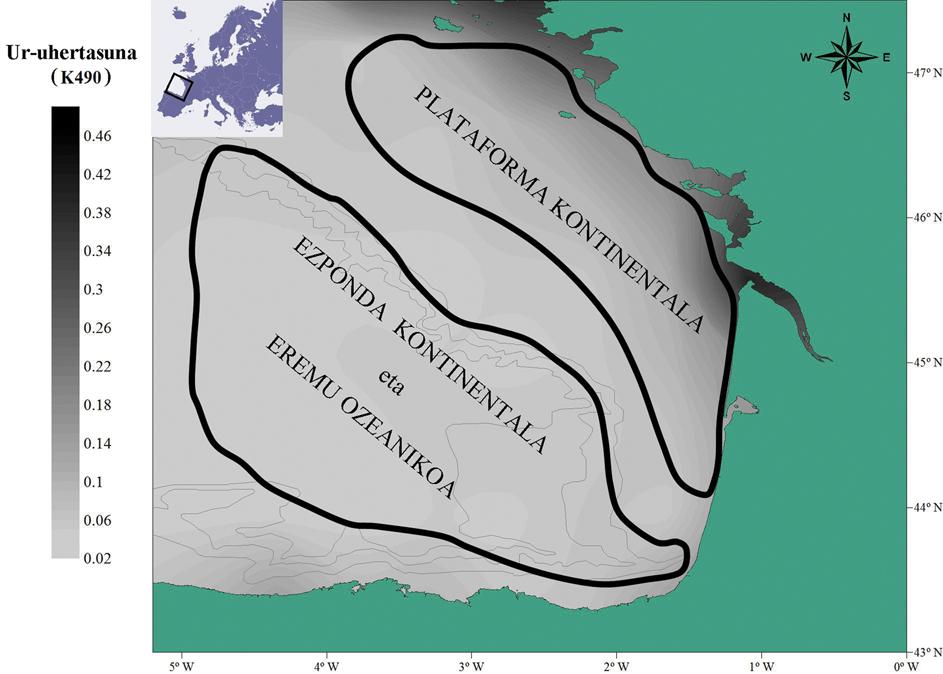
Pero, ¿cómo podrían ser las dos primeras zonas mencionadas, a pesar de tener menos alimentos? Por un lado, porque al no ser tan turbulento el agua de la zona, se puede atrapar más fácilmente el gran zooplancton que parece más interesante energéticamente, y por otro, porque aumenta la posibilidad de escapar a los jóvenes de anchoa de las presas. Además, parece que el alto grado de llenado gástrico (elevado peso) puede ser un indicador de buen estado biológico, lo que puede contribuir en gran medida a la supervivencia invernal (reclutamiento) del stock juvenil de anchoa.
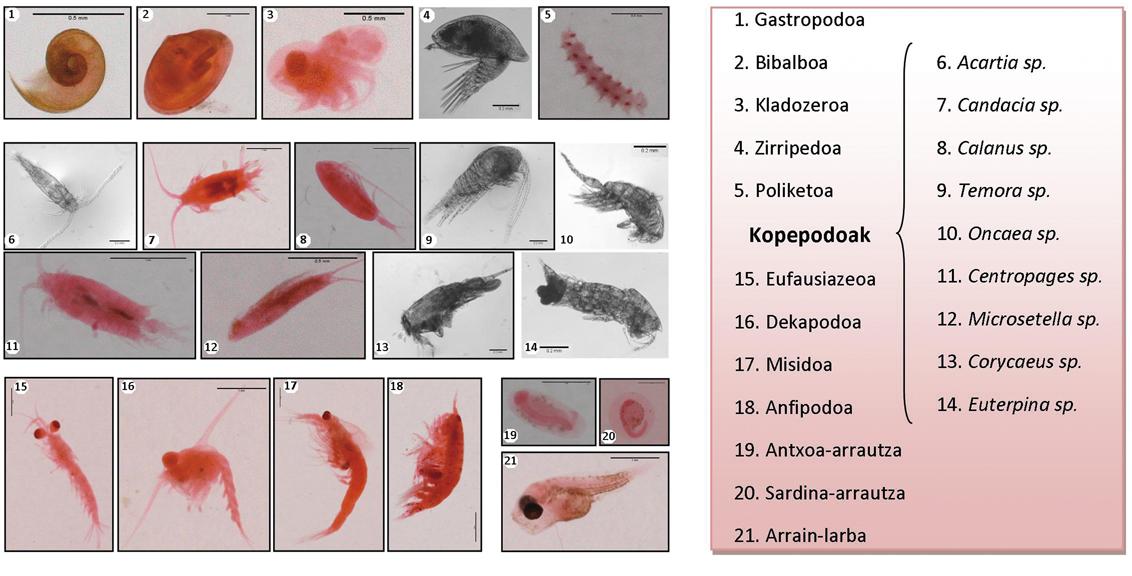
Sin embargo, la información que nos aporta el peso de los estómagos es útil pero, por otra parte, limitada, por la apertura del estómago de los peces y por las diferencias que se han observado entre especies es de especial interés. Analicemos, por tanto, cómo son las dietas de las principales especies pelágicas consideradas en las campañas oceanográficas de investigación de los años 2008 y 2009 (BIOMAN, ECOANCHOA, PELACUS, JUVENA) de mayo a septiembre y durante el día y la noche en las distintas zonas geográficas del golfo de Bizkaia.
Capturas de peces: muchos pequeños, pocos grandes
Todo lo que los peces tienen a su disposición en el mar, si es accesible por vía oral, ¿lo comen? Sí. Esta pregunta parece sencilla. Pero, ¿qué sugeriríamos si dijéramos que los peces grandes comen principalmente presas pequeñas? De hecho, las anchoas del golfo de Bizkaia ( E. encrasicolus ), las sardinas ( S. pilchardus ), los chicharros negros o atlánticos ( T. trachurus ), los chicharros blancos o mediterráneos ( T. mediterrus ), el verdel atlántico ( S. scorrelus ), los chopeles ( Bogs. colias ), el entorno de la competencia para la comida ( Sellés ( Ratos ( Ratos ) y Ratos ( ). Por otra parte, tanto el tamaño de las capturas como de las capturas pueden determinar en parte su biología y ecología. En este sentido, los resultados de este estudio han demostrado que un alto ancho de boca respecto al tamaño corporal no tiene por qué limitar el tamaño de la presa/presa ni la distribución del ratio de biomasa. De acuerdo con ello, los depredadores capturan a las presas de todo el rango de tamaños disponibles, incluidas las de menor tamaño, y, a medida que van creciendo, añaden mayores presas a su dieta, siempre que puedan ser capturadas. Se observa, por tanto, un alto grado de solapamiento en la composición de la dieta, incluso entre peces de distinto tamaño. Así, los chicharros son los únicos que parecen especializados en grandes presas.

El menú de pescados no depende de su tamaño
Las diferencias entre las dietas de los peces pequeños pelágicos también pueden estudiarse atendiendo a las diferencias en el comportamiento alimentario asociado a la posibilidad de obtener presas. Se ha comparado el zooplancton de cada punto de muestreo (muestras de agua tomadas con las redes de Nozovet) con el contenido de estómago de los peces autóctonos, y los resultados indican que los peces que comparten una misma fosa se alimentan de la misma fuente de alimentos y que su alimentación está compuesta mayoritariamente por pequeños zooplancton (por ejemplo, copépodos). De hecho, los chicharros y verdel más grandes, por ejemplo, comen las mismas especies de presa que las anchoas más pequeñas. En cuanto al consumo de grandes presas (por ejemplo, eufausiáceos, decápodos y misidos) que requieren mayor esfuerzo para los peces, este depredador se ha observado con mayor frecuencia en los casos de chicharro negro y blanco, así como en los casos de verdel atlántico y makaela. Por lo tanto, podríamos decir que las dos especies mencionadas parecen dominar al resto, como la anchoa, la sardina, la boga y la sardina de gitano, a medida que muestran una mayor tendencia a la captura de grandes presas, sobre todo durante la noche.
Además, como se ha visto en el caso de las anchoas jóvenes, estos resultados sugieren que la limitación visual puede jugar un papel importante en el éxito de los peces en la depredación. No obstante, dado que la dieta está compuesta principalmente por organismos de pequeño tamaño relativo, las dietas son similares en cuanto a las especies ingeridas. Sin embargo, atendiendo a la composición de la dieta, se puede apreciar un elevado grado de solapamiento entre pequeños peces pelágicos, que varía en función de la especie, tamaño, tiempo y zona geográfica de la presa. Por otro lado, parece que el zooplancton disponible en el mar limita la diversidad de las capturas ingeridas, más que las preferencias de los peces hacia las presas. De las ocho especies investigadas, la mayor superposición de la dieta se da entre las tres especies de clupeidos (anchoa, sardina y sardina de gitano), y las anchoas presentan una mayor actividad alimentaria que las sardinas gitanas respecto a las presas relativamente mayores. Sin embargo, el solapamiento dietético no tiene por qué implicar competencia alimentaria, al menos mientras sobra alimentación, especialmente si hablamos de pequeños zooplancton (por ejemplo, copépodos).

Influencia ecológica de la cadena trófica en las anchoas
Lo expuesto hasta ahora puede poner en duda la utilidad de esta investigación para la gestión de la pesca, ya que el simple conocimiento de lo que comen los peces no parece que sirva para garantizar la sostenibilidad de la población piscícola. Sin embargo, las interacciones entre los depredadores y, en particular, entre los depredadores, son en la actualidad el tema principal de la investigación pesquera, ya que el conocimiento de la dinámica de las poblaciones de peces puede garantizar la eficacia y sostenibilidad de la futura pesca. ¿Por qué? Vamos a verlo.
El zooplancton, los huevos de peces y las larvas prematuras de los peces pelágicos son planctónicas y, en consecuencia, potenciales presas para las especies piscícolas planctivas que compiten con sus padres por adquirir alimento (lo que denominamos "depredación entre competidores"). Tanto el canibalismo como la depredación entre competidores han sido considerados por estudios previos como mecanismos de regulación entre especies en competencia. Teniendo en cuenta esto, se ha estimado la tasa potencial de mortalidad que supone la depredación de las principales especies pelágicas menores (incluida la depredación de las anchoas, que puede considerarse canibalismo) en los tempranos estadios de la anchoa, es decir, en los huevos y larvas de las anchoas. El muestreo para este estudio se realizó durante la puesta de la anchoa, por lo que en el estómago de los depredadores no se encontró apenas larín. Sin embargo, atendiendo al número de huevos de anchoa ingeridos por diferentes especies de peces, los resultados indican que la depredación de los clupeidos y, en especial, de las sardinas, puede tener una gran influencia sobre la supervivencia de los huevos de anchoa: el impacto de la sardina y el verdel es superior al que puede causar el chicharro negro, el chicharro blanco, el bogas, la sardina de gitano o la anchoa.
Así, los resultados de 2008 sugieren, por ejemplo, que cerca del 33% de los huevos de anchoa se perdieron por depredación de la sardina, valor que se situaba en el 37% si se tenía en cuenta la depredación de todas las especies pelágicas pequeñas antes mencionadas. También se observó que estos valores pueden variar mucho año tras año. La frecuencia del canibalismo y de la depredación entre competidores puede variar en función de la abundancia relativa de las especies de predadores y presas, su concentración y la superficie de solapamiento del hábitat. Por lo tanto, podríamos decir que las investigaciones que se realizan para estudiar el reclutamiento de la anchoa y asesorar sobre el stock deberían tener en cuenta la depredación de los huevos de anchoa, ya que ahora sabemos que la alimentación de las especies pelágicas influye directamente en la mortalidad de la anchoa en su fase temprana y por tanto en su supervivencia.

Bibliografía
Zu idazle
Zientzia aldizkaria

