

Orgánulos sin membrana
Cuando en las escuelas de biología celular se explican las células y sus características, siempre se mencionan dos tipos principales de células: la célula eucariota y la célula procariota. Y por otro lado, cuando se enumeran las diferencias entre ambos tipos de células, antes o después aparecen los orgánulos. Los orgánulos suelen definirse como estructuras especializadas que cumplen funciones específicas en el interior de la célula. En las explicaciones sobre los orgánulos siempre se ha mencionado el núcleo, protector de la información genética celular; las mitocondrias y cloroplastos, productores de energía; el retículo endoplasmático granulado y liso, que participan en la síntesis de lípidos y proteínas; y el aparato Golgi, compuesto por vesículas de secreción que forman y distribuyen paquetes de proteínas. Todas ellas tienen una función específica y están rodeadas de una membrana semipermeable.
En los últimos 100 años no ha habido cambios en la estructura celular que conocemos. ¿O sí? En noviembre de 2018, investigadores del Instituto Sloan Kettering publicaron un nuevo órgano en la revista Cell (Ma & Mayr, 2018). ¡Nuevo orgánulo asignado a un dominio TIGER (que explicaremos más adelante)! ¿Seríamos ante un nuevo concepto que cambiara la integridad de la célula?
En los últimos años, los investigadores han analizado con más detenimiento la sopa de agua y moléculas tanto del citoplasma celular como del interior de los orgánulos. Los investigadores han observado en el citoplasma y en el interior de los orgánulos (es decir, en el interior de los orgánulos con una densidad determinada rodeados de líquidos de otra densidad) una gran cantidad de vesículas sólidas que, al igual que los orgánulos membranosos, tienen funciones especializadas. Estas vesículas a menudo no se corresponden con la definición de un orgánulo “clásico”: por un lado, estos orgánulos no membranosos no tienen límites concretos, y por otro, pueden ser orgánulos efímeros, pueden reaccionar rápidamente bajo la influencia de las señales químicas y, al mismo tiempo, desorganizar estructuras especializadas generadas en la desaparición del estímulo.
Sin embargo, por su función e importancia han conseguido la atención de los investigadores.
Hay varios orgánulos sin membrana
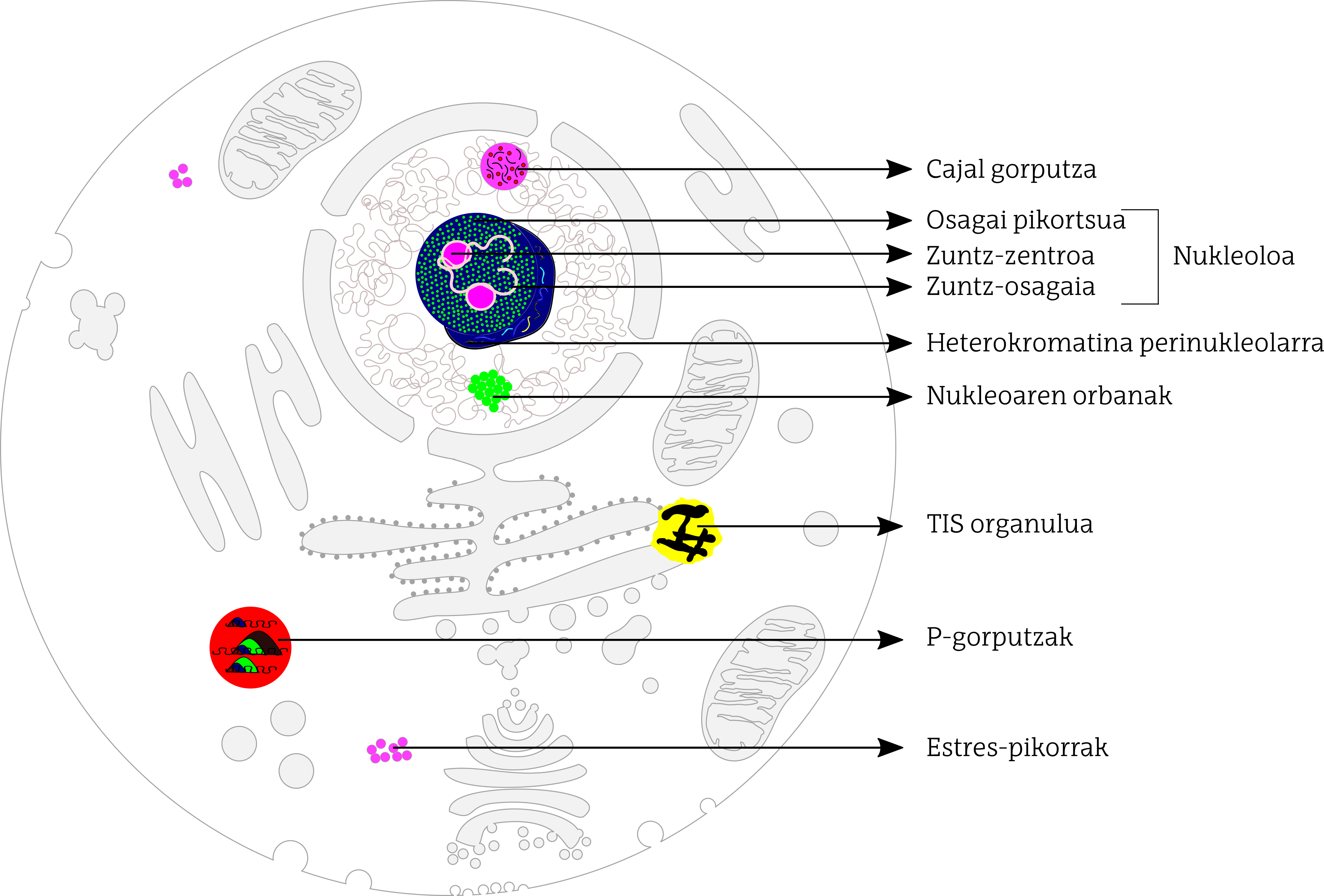
En las células de los mamíferos, los investigadores han identificado al menos una docena de orgánulos sin membrana dispersos tanto en el núcleo celular como en el citoplasma (Figura 1). En general, los orgánulos no membranosos que se encuentran dentro del núcleo están especializados en el control de la transcripción de genes y en algunos aspectos del metabolismo del ARN. Sin embargo, los orgánulos no membranosos dispersos por el citoplasma intervienen en el metabolismo, transporte y homeostasis del ARN mensajero.
a) Orgánulos sin membrana dentro del núcleo
Entre los orgánulos no membranosos que se encuentran dentro del núcleo, destaca el nucleolo. El nucleolo es el más estudiado y probablemente el más grande de todos. Su función principal es la biogénesis de los ribosomas necesarios para sintetizar proteínas. En el nucleolo se distinguen tres regiones: centro de fibras, componente fibroso denso y componente granular. En la primera se sitúa el ADN que codifica los ribosomas; la conversión al ARN de este ADN, la transcripción, se produce en el límite entre el centro de fibra y el componente de fibra. En este límite aparece la proteína conocida como fibrilarina y se produce el procesamiento del ARN que codifica los ribosomas, la adhesión al corte. El componente granular, por su parte, es rico en proteínas nucleofosmina, donde se produce la acumulación y ensamblaje de subunidades prerribosómicas. El nucleolo está rodeado de ADN organizado de forma muy compacta, a la que se denomina heterocromatina perinucleolar. Son principalmente ricos en repeticiones de ADN, principalmente satélites de ADN y conjuntos de ADN ribosómicos. Parece que esta repetición del ADN del ribosoma evita la reconversión o recombinación del ADN, estabilizando el ADN ribosómico.
Los cuerpos de Cajal también se encuentran en el núcleo de la célula, pero son más pequeños. En cuanto a la estructura, están constituidos por proteínas y RNA que parecen ser necesarios para el ensamblaje y modificación de snRNP (small nuclear ribonucleoprotein). Estos snrnp son necesarios para los cambios posteriores a la transcripción del RNA mensajero: entre otros, las partes transcritas de RNA que no se convertirán en proteínas, las intronas, las adhesiones de corte. Los cuerpos de Cajal parecen participar en el proceso de maduración final de las partículas snRNP y en la generación de complejos snRNP.
Las manchas del núcleo están directamente relacionadas con los cuerpos de Cajal y forman parte del nucleoplasma. En las manchas del núcleo se acumulan y modifican snRNP y proteínas ricas en aminoácidos serina y arginina. Todos ellos participan en el proceso de corte y pegado del ARN mensajero.
b) Orgánulos sin membrana dispersos en citoplasma
En cuanto a los orgánulos no membranosos dispersos por el citoplasma, generalmente se denominan gránulos mRNP (messenger ribonucleoprotein). Aunque existen varios tipos de RNP, es habitual que las proteínas y el ARN mensajero se compartan e interactúen.
Los cuerpos de P (processing bodies) se han descrito en diferentes tipos de células y se ha observado que son áreas ricas en proteínas que intervienen en el transporte, modificación y traducción del ARN mensajero. Cuando se acumulan muchos ARN mensajeros no traducidos a aminoácidos, bien porque la traducción está inhibida o en ciertas condiciones de estrés, aumenta el tamaño y el número de cuerpos P.
Los gránulos de estrés, como su nombre sugiere, se ensamblan ante señales de estrés, secuestrando moléculas de RNA mensajero silenciadas y factores de traducción. Es decir, la célula almacena en zonas concretas el material necesario para hacer frente a este estrés. En los gránulos de estrés es habitual encontrar los factores necesarios para iniciar la traducción y los componentes de las subunidades pequeñas de los ribosomas.
Otros orgánulos citoplasmáticos sin membrana son los gránulos germinales, que aparecen en las células germinales del embrión en desarrollo (células madre creadoras de células sexuales). Normalmente son ricos en ARN mensajero y en enzimas que modifican el ARN. Parece que intervienen en los cambios posteriores a la traducción del ARN mensajero en las células de la línea germinal.
En septiembre de 2018 publicaron en la revista Cell el descubrimiento del último tipo de orgánulos sin membrana: Fue conocido como el orgánulo TIS. En interacción con el retículo endoplasmático, permite interacciones entre proteínas dirigidas por el extremo 3’UTR del ARN. La proteína TIS11B está muy relacionada con este orgánulo, de ahí su nombre. La interacción entre el orgánulo TIS y el retículo se denomina dominio TIGER. Este último es diferente al resto de medios del citoplasma, tanto biofísicos como bioquímicos. Todo ello controlará el intercambio de proteínas del retículo endoplasmático, regulando las interacciones entre proteínas, relacionando los extremos 3’UTR con la proteína TIS11B y organizando una red en el citoplasma y alrededor del retículo liso.

Relacionadas con enfermedades
El alto dinamismo de los orgánulos no membranosos hace que no sea fácil establecer relaciones directas entre los problemas de funcionalidad y las enfermedades de estos orgánulos. No obstante, cabe destacar el caso de la esclerosis lateral amiotrófica. Una de las características de esta enfermedad (como otras enfermedades neurodegenerativas) es que se producen inclusiones de mensajeros de ribonucleoproteínas (RNP) que alteran el metabolismo normal del ARN mensajero (Figura 2). Estas inclusiones patológicas están formadas por gránulos proteicos RNP, cuya gran acumulación produce un ensamblaje entre ellos y la formación de fibras amiloides. Las fibras amiloides alteran el funcionamiento normal de las células, degradándolas.
En los casos de cáncer, los orgánulos no membranosos también presentan interesantes opciones o perspectivas. Uno de los factores que rigen el crecimiento celular es el ensamblaje de ribosomas, que como se ha visto tiene una gran importancia en la producción de ribosomas. El nucleolo, por tanto, puede ser un importante destino de los tratamientos contra el cáncer, caracterizado por el crecimiento incontrolado del nucleolo y el aumento del nucleolo.
¿La clave del nacimiento de la vida? Repensando Oparin
En 1922, el bioquímico ruso Alexander Oparin publicó una teoría sobre el origen de la vida en los inicios de la historia de la Tierra. Según su teoría, se formaron los primeros compuestos orgánicos simples, ya que la energía eléctrica de los rayos o la energía calorífica de los volcanes provocó la reacción del metano, el amonio, el agua y otros componentes, ricos en la atmósfera reductora de la época. Todos estos ingredientes se organizaron en pequeñas gotas denominadas coacerbatos. En el año 2016 un grupo de investigadores alemanes describieron los activos químicamente y las gotas de líquido con capacidad de fragmentación y, por tanto, de perpetuación. Estas gotas y los orgánulos no membranosos tienen algunas características iguales, por lo que ¿existirán claves para explicar el origen de la vida en los orgánulos sin membrana?
Referencias
Crabtree, M., Nott, T. 2018. These organelles have no membrane. The Scientist. Diciembre 2018. https://www.the-scientist.com/infographics/infographic--what-are-membraneless-organelles--65135
Gómez, E., Shorter, J. 2018. "The molecular language of membraneless organelles". Journal of Biological Chemistry 1-23.
Ma, W., Mayr, C. A. 2018. "Membraneless Organelle Associated with the Endoplasmic Reticulum Enables 3< UTR-Mediated Protein-Protein Interactions". Cell 175: 1492-1506.
Mao, Y.S., Zhang, B., Spector, D.L. 2011. 'Biogénesis and Function of Nuclear Bodies'. Trends in Genetics 27: 295-306.
Mitrea, D.M., Kriwacki, R.W. 2016 "Phase separation in biology; functional organization of a higher order". Cell Communication and Signalling 14: 1.
Zu idazle
Zientzia aldizkaria

