

El remolino de los últimos mohicanos: los peligros de la escasez

El ser humano, con una enorme capacidad de transformación del entorno, ha colonizado el mundo entero transformando el hábitat de muchos seres vivos y sobreexplotando muchos recursos. Como consecuencia de ello, la población se ha reducido considerablemente creando un conjunto de problemas que la impulsarán a la extinción.
Las poblaciones pequeñas tienden a desaparecer y en ello influyen diversos factores. Entre ellos cabe destacar:
• Variabilidad demográfica
Las poblaciones pequeñas siempre dependen del azar, en el límite de la extinción. En muchas especies es posible cuantificar el número medio de crías, la mortalidad media o la proporción de machos y hembras en recién nacidos. Si el tamaño de la población es grande, estos cálculos pueden ser bastante precisos. En poblaciones pequeñas, sin embargo, el azar tiene una gran influencia. Por ejemplo, en la mayoría de las especies, la mitad de los recién nacidos son machos y la otra mitad son hembras. Pero la mitad de las crías de una hembra no deben ser necesariamente hembras. Así, si la población es muy pequeña, puede ocurrir que haya pocos recién nacidos, que se desvíe mucho la distribución sexual o que mueran muchos ejemplares en un momento determinado. A diferencia de las poblaciones grandes, en las pequeñas la extinción siempre está cerca y si por cualquier motivo la cantidad baja a cero, la población no podrá recuperarse.
Estas fluctuaciones inducidas por el azar se denominan estocasticidad demográfica. Supongamos una población de pájaros formada por dos parejas con un promedio de 3 hijos. La población debería aumentar, aunque la probabilidad de crecimiento de la población en la siguiente generación es de 20/64, de mantener 30/64, de descender 12/64 y de desaparecer 2/64 (ver tabla 1).
Posibles
combinaciones de descendientes
Macho Macho Macho Macho Macho Macho Macho Macho Macho Macho Macho Macho Hembra Hembra Macho Hembra Hembra Hembra
Macho
Hembra Hembra Hembra Hembra
6/64
15/64
20/64
15/64
6/64
1/640 1
2
2
1
0Destrucción Mantener Incremento Mantener Reducción Destrucción
• Problemas genéticos

Como se ha comentado anteriormente, la variabilidad demográfica es un problema grave en poblaciones pequeñas, pero no el único. Además, las poblaciones pequeñas presentan importantes problemas genéticos. La información genética de estos seres vivos está dividida en unidades denominadas genes y la localización de cada gen en el cromosoma se denomina locus. En cada locus pueden existir distintas formas génicas, alelos. La diversidad genética existente en la población, es decir, la diversidad de alelos, es muy importante, ya que permite adaptarse a los cambios del medio.
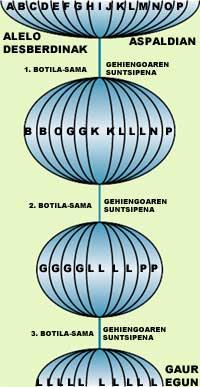
Cuando los individuos se cruzan para reproducirse, sus genes se mezclan aleatoriamente y, por tanto, en las generaciones futuras aparecen nuevas combinaciones genéticas. Si un alelo es escaso en poblaciones pequeñas, puede desaparecer fácilmente al azar y, por tanto, disminuir la diversidad genética. La variación del grupo de genes que puede producirse aleatoriamente se denomina deriva genética (Tabla 2, Figura 1). Su influencia es aún más acusada en poblaciones pequeñas y, sobre todo, si en un momento dado esa población ha pasado por un número mínimo de ejemplares (una botella sama). Por ejemplo, en el cráter de Ngorón de Tantzania vivían entre 60 y 75 leones hasta que en 1962 proliferaron los parásitos y murieron muchos leones. La diversidad genética se redujo considerablemente y a pesar de que en 1964 se introdujeron nuevos ejemplares, hoy en día la diversidad genética es muy baja y se observan numerosas anomalías en el esperman (Figura 2).
AA= 1,8,50 AAA
= 0,87,8 AAA
= 2,85,7AAa; 0,5AA2,5U0,0,0,0,0,H
= 2,5A0,0,0,A; 0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,3,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,0,
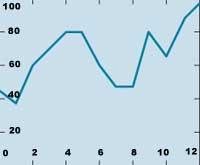
En poblaciones pequeñas, la diversidad genética puede perderse por otra vía: debido a la abundancia de cruces entre sus componentes sanguíneos, aumenta la probabilidad de que en un gen aparezca el mismo alelo, con la consiguiente pérdida de heterocicencia. Según Lacy, las poblaciones de quinientos individuos a cien generaciones mantienen el 90% de su diversidad original, mientras que las de veinte individuos sólo lo hacen el 5% (ver figura 3).
La pérdida de variabilidad puede ser compensada por la mutación y la inmigración, pero la recuperación de las bajas tasas de mutación que se dan en la naturaleza no es fácil: el cruce con los ejemplares de otro grupo puede producir una enorme variabilidad genética.
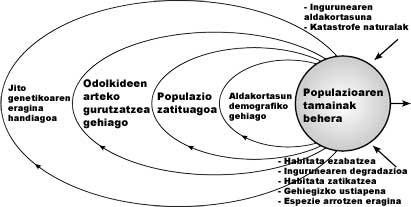
• Alteraciones del medio y catástrofes naturales
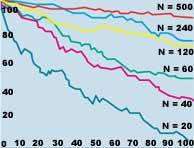
Los cambios aleatorios del medio se denominan estocasticidad ambiental. Los cambios ambientales afectan a poblaciones de todas las dimensiones, pero, por supuesto, su tamaño tiene mucho que ver. Por ejemplo, cuando un rayo mata un grano en una población de quinientos ejemplares, la dinámica propia de la población no varía significativamente, mientras que si esta población está compuesta por sólo diez individuos, el efecto es mucho más violento. Este tipo de estocasticidad afecta más directamente que la demografía a la viabilidad de la población (ver figura 4).
De lo anterior se concluye que cuanto menor sea la población, mayor será la influencia de la variabilidad ambiental, demográfica y de los problemas genéticos, lo que redundará en un mayor riesgo de destrucción. Este fenómeno se denomina Efecto Zurrunbilo. Por tanto, la disminución de la población en la disminución del hábitat se traducirá en una mayor influencia de la variabilidad demográfica, con una disminución del tamaño, al tiempo que aumentará la incidencia de la deriva genética y una vez introducida en este remolino, la extinción de la población puede considerarse prácticamente segura (ver figura 5).
Análisis del estado de la población
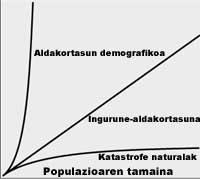
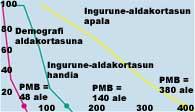
Si conocemos la variabilidad del medio, la variabilidad demográfica de la población y la diversidad genética, podemos calcular cuál es la población más pequeña que puede permanecer sin riesgos o hasta qué punto es viable una población de cierta dimensión. La primera se denomina Población Mínima Viable (PMP) y la segunda Análisis de Viabilidad de la Población (PBA). Shafer definió la PMB como la menor población aislada con un 99% de probabilidad de subsistencia en 100 años, teniendo en cuenta la estocasticidad demográfica, ambiental, genética y catástrofes naturales.
Para ello, por supuesto, necesitamos mucha información (fisiología, etología, genética, distribución, autoecología, dinámica del medio y de la población, etc.) y en muchos casos la mayoría de estos datos son desconocidos. Por ejemplo, Lande propuso la represión de 1.000 ejemplares de vertebrados, que en el caso de los invertebrados, debido a la variabilidad de las poblaciones, obligó a 10.000 ejemplares. Sin embargo, mantener una población de miles de individuos no es fácil, sobre todo en el caso de los grandes vertebrados, cuya Demanda de Superficie Mínima (RME) es muy elevada. Según la estimación de Belovsky, los grandes mamíferos necesitarían reservas de entre 100.000 y 1.000.000 km 2 para sobrevivir en un siglo al 95%. Si tenemos en cuenta que la superficie de las reservas mundiales es de 4.846.300 km 2, la protección de la diversidad de especies implica necesariamente la extensión de los espacios protegidos.
Categoría Mace LandeMaceEnestado
crítico Amenazados Vulnerables 50% 5 años o
2 generaciones 20%
20 años o 10% 100 años en
10 generaciones 50%
5 años o
2 generaciones 20%
20 años o
5 generaciones 10%
50 años o
10 generaciones.
El índice PBA también puede utilizarse como criterio de clasificación en el libro rojo del estado de conservación de las especies publicado por la IUCN. En función de los resultados del PBA se propone una clasificación con tres categorías: especies vulnerables, especies en peligro y en situación crítica (ver tabla 3). El problema es que no todas las especies son conocidas al mismo nivel y que en la cuantificación de los parámetros se requiere mucha precisión. Para analizar la influencia de los fenómenos estocásticos mencionados en el riesgo de extinción se realizan modelos de simulación por ordenador que permiten medir el PMB (ver figura 6).
Los modelos construidos hasta el momento se han utilizado para poblaciones aisladas. Pero en la naturaleza, las poblaciones están divididas en un mosaico llamado metapopulación. Como ya se ha mencionado anteriormente, la migración entre ellas, además de aumentar la variabilidad genética, actúa de otras formas en la viabilidad de la población: al cambiar las condiciones de un lugar a otro, no todas sufrirán alteraciones catastróficas o ambientales y, por tanto, a pesar de la desaparición de una población, otra puede recolonizar el territorio dejado por ella, permitiendo así la duración de la metapopulación. Por ello, sería conveniente construir un modelo que contemple las interacciones entre poblaciones (Figura 7).
Un ejemplo de ello es el del oso pardo de las Montañas Rocosas. Según las estimaciones realizadas por Shafer en 1983, la PMB es de 50 ejemplares, mientras que Bolger la estima de 220. El AME requerido por esta cantidad sería superior a la mayor reserva de EEUU. Si sólo quedan seis osos en el Pirineo, su probabilidad de supervivencia sería menor de 20 años1 (ver tabla 4).
Número de poblaciones100 años de probabilidad (%)Duración más probable(años)10 20
30
40
506
36
74
94
9819
44
69
79
114
Dado que los osos pirenaicos no pueden perdurar en el futuro, la gestión de los osos debería realizarse a nivel de metapopulación, incluyendo las poblaciones fuera de este ámbito. De este modo, la introducción de osos procedentes del exterior permitiría una mayor viabilidad de los osos pirenaicos. Por otro lado, aunque se cree que la población de lobos ha aumentado, en la península Ibérica su futuro es bastante incierto, ya que en la actualidad sólo existen 500 ejemplares, la mitad de los 1.000 ejemplares propuestos por Land.
Mirando al futuro
Las sesiones de cálculo de PBA, PMB y AME llevadas a cabo en los últimos años nos han hecho ver una imagen preocupante. Muchas poblaciones no serán viables a largo plazo, ya que se necesitan más ejemplares de lo que se pensaba para asegurar su supervivencia y las zonas de protección que se necesitarían son mucho más amplias que las actuales. Pero no hay casos sin esperanza, sino gente poco esperanzada y casos caros. En este sentido, se constata la necesidad de corregir dos problemas: la necesidad de reservas más amplias y poblaciones mayores, lo que nos permitiría sentar las bases para trabajar en la conservación de las especies. Posteriormente, se debería realizar el trabajo correspondiente, pudiendo incluirse en las medidas y planificaciones requeridas por la conservación para cada especie y contribuir, lógicamente, a huir del remolino del último mohicano.
Zu idazle
Zientzia aldizkaria

