

Nuevos avances en biología XIX. siglo
Como hemos visto antes, XIX. Para finales del siglo XX los amantes de la naturaleza consideraban definitiva la obra de Darwin, ya que la evolución a través de la selección natural era considerada como un principio establecido. A continuación, la investigación embriológica para lograr una mayor precisión la veían para caminos prósperos, ya que según las hipótesis de Meckel y Haeckel, la historia de un ser embriológico sería rehacer en miniatura la historia de su especie.
Podemos pensar que había excepciones. Por un lado, De Vries investigaba sobre los cambios y alcanzó un enorme prestigio por sus consecuencias hereditarias. Por otra parte, Bateson en 1890 criticó las pruebas de la “ley” de Haeckel y sus fundamentos lógicos y propuso volver a los caminos de Darwin. Las ideas de Darwin sobre los orígenes de las especies que entonces se encontraban en la pila, tuvieron algunos obstáculos. Las más importantes fueron, según Bateson:

“La primera dificultad se refiere a la magnitud de los cambios que dan lugar a nuevas formas. Se considera que los cambios que producen las especies en todos los seres antiguos sobre la evolución son pequeños, aunque muchas veces no se diga explícitamente. Pero si son pequeños, ¿qué ventajas o ventajas aportan a los que los tienen sobre sus compañeros? Esta dificultad se conoce con el nombre de “pequeños cambios o iniciales”.
La segunda dificultad es similar. Considerando que estos cambios se producen, si se mantienen y se perpetúan, generarían nuevas especies. ¿Cómo los pueden mantener? ¿No se pueden eliminar estos cambios cuando se mezclan con los que no tienen seres que tienen? A esta segunda dificultad se le denomina "resultado destructivo del cruce".
A esto Bateson añadió que, como sabe cualquier persona que se dedica a la cría de plantas o animales, si bien en muchos casos aparecen pequeñas variaciones con respecto a lo que se considera normal en los seres, en otros muchos son grandes. Las investigaciones sobre este tema para finales de siglo bastaban para comprobar que las mutaciones bruscas no eran muy ocasionales y que muchas veces estas mutaciones pasaban a sus descendientes. De esta forma pueden aparecer nuevos cambios, pero no especies.
Sin embargo, el motivo y la causa de estos cambios no se conocían y, tal y como aparecían, debían tomarse simplemente. Pero en aquellos años se conocieron nuevas obras, mejor dicho, se olvidaban de antiguas obras de interés como las de Mendel.
Johann Mendel nació en Henomorf (Alemania) en 1822 en una familia campesina. Tras los estudios elementales, entró en el convento de los agustinos y a los veintiún años se ordenó sacerdote, pasando posteriormente a denominarse Gregor, el mundo de la ciencia. Su deseo de dedicarse a la enseñanza de la ciencia le llevó a realizar estudios especiales en la universidad de Viena. Luego en la Escuela Superior de Br\nn enseñó a lo largo de catorce años física y ciencias naturales. Sin embargo, realizó su mayor trabajo y sus investigaciones más prolíficas en el propio monasterio de Brünn.
En 1856 Mendel inició sus investigaciones cruzando guisantes de distintas razas en la huerta de su monasterio. Aunque al principio parece que lo consideró como un pasatiempo o un juego, pronto, a la vista de los resultados, se desencadenó una pasión investigadora con personalidad propia. Con ensayos repetidos y cambios descubrió las leyes generales de la hibridación. Tras ello, durante ocho años, en solitario, se dedicó a comprobar los logros obtenidos. Durante este tiempo, como se dice, exploró minuciosamente doce mil plantas.
Todo este trabajo lo recogió en el informe titulado “Ensayos sobre plantas híbridas” y lo explicó a la Asociación de Historia Natural de Brognn en 1865. Aquella asociación, al parecer, no era de las más altas y no le dieron ninguna importancia.
A la vista de ello, Mendel envió su obra a unos científicos (entre otros, Von Nágeli, catedrático de la botánica de Munich, muy alto en aquella época), pero de él sólo recibió una respuesta de cortesía, quizá por considerarla como consecuencia de un juego de entretenimiento de un fraile botánico.
Ante la mala acogida que sufrió, y viendo la dificultad para aprobar sus obras, Mendel perdió poco a poco su pasión por la ciencia. En 1868 fue nombrado prelado y debido a los trabajos que este cargo le generó en la diócesis, dedicó sus esfuerzos a la ciencia en este sentido. Por ello, hasta su muerte, el 6 de enero de 1884. Se dedicó a las tareas eclesiásticas.
En el informe titulado “Ensayos sobre plantas híbridas” se recoge una declaración de las leyes que sustentan la genética actual y que llevan el nombre de Mendel. En la introducción del trabajo, Mendel recordó los trabajos de los científicos que antes había trabajado en las investigaciones de los híbridos, y explicó en qué consiste y cómo abordará el problema.
En este trabajo aparecen dos leyes fundamentales: la primera es la de la disfunción de caracteres (que ya se conocía anteriormente) y la segunda la de la independencia de dichos caracteres. La investigación de Mendel comenzó explorando un guisante, el Pisum sativum. Estudió el cruce de las guisantes de semilla lisa (L) y rugosa (Z). En la primera generación, llamada F1, sólo aparecen semillas suaves. Pero si estos últimos se cruzan entre sí, en la segunda generación (F2) aparecen el 75% de guisantes tipo L y el 25% tipo Z. En el híbrido F1 el carácter Z (subyacente) estaba oculto y sólo aparecía el carácter L (principal), pero en el híbrido F2 volverá a aparecer Z.
Las leyes de Mendel lo permiten. El híbrido F1 es de tipo LZ, donde los gametos L y Z son iguales. Por ello, una vez cruzados estos híbridos, en el híbrido F2, tendremos los siguientes cuatro tipos de híbridos: LT, LZ, LZ y ZZ. En ellos los de LL serán suaves. También los de LZ, ya que el carácter L es el principal respecto a Z, pero los de ZZ serán rugosos. Por lo tanto, las proporciones anteriores serán del 75% lisas y del 25% rugosas.
Mendel también utilizó guisantes con varios caracteres: semillas lisas y amarillas por un lado y arrugas y verdes por otro. En este caso existían cuatro tipos de gametos: pulido amarillo-rugoso, rugoso-amarillento, pulido-verde y rugoso verde, por lo que dieciséis combinaciones en las proporciones 9, 3, 3, 1, siendo lisa y amarilla el carácter principal y verde y la rugosidad inferior.

Debido a los resultados obtenidos por Mendel, se observaba que los caracteres hereditarios estaban relacionados con los elementos que podían ser distribuidos, por lo que se indicaba una discontinuidad en el patrimonio de la herencia.
En Mendel aparecen todos los elementos de la genética moderna: las leyes de la hibridación y sus usos. La conceptuación de Mendel fue muy concreta. No se trata de una intuición nublada como ocurre en los trabajos iniciales, sino de algo maduro y humillado. Dejando a un lado la terminología, la obra de Mendel puede considerarse hoy escrita. Por lo tanto, en el campo de la ciencia de la herencia, Mendel no puede ser considerado un pionero, sino un creador de dicha ciencia.
Como hemos dicho antes, todo este trabajo fue descartado y aquel fraile, con una gran paciencia, tuvo que dejar todos sus logros a los treinta y cinco años de silencio, es decir, hasta su muerte. En 1900 De Vries, Correns y Tscherma descubrieron, confirmaron y divulgaron los trabajos de Mendel.
Debido a la tendencia aceptada en el mundo de la física actual, resulta muy adecuado expresar las cualidades biológicas, por decirlo de alguna manera, a través de unidades atómicas bajo la legislación de probabilidad. Si no es posible predecir el movimiento de un átomo o electrón concreto, ocurre lo mismo con el tipo de herencia que aparecerá en un organismo especial, pero en ambos casos podemos calcular probabilidades y si tomamos un número muy elevado podemos tener seguridad de que se cumplirán las predicciones.
Como hemos visto, en la investigación de la herencia hay que tener en cuenta la especificidad de los caracteres, es decir, si son predominantes o sub-decisivos. Uno puede transmitir el carácter principal a sus descendientes, siempre y cuando tenga claro que lo tiene.
Por el contrario, en algunos casos puede aparecer algún carácter subyacente sin que previamente se haya dado cuenta en alguno de los siguientes. En el caso de que dos seres que llevan un carácter subyacente (aunque no aparezcan) en sus células se fecundan entre sí, dicho carácter subyacente aparecerá claramente en el 25% de sus descendientes.
Sin embargo, en la mayoría de los casos los problemas hereditarios son mucho más complicados que lo visto con dos caracteres. Además, algunas cualidades pueden ser predominantes o sub-sexuales. Otras veces los caracteres aparecen por parejas, es decir, uno no aparece si no aparece el otro, y viceversa. En otros casos son incompatibles y nunca aparecerán juntos.
Muchos caracteres mendelianos se han identificado en plantas y animales y se ha utilizado este método para mejorar las razas, aumentar ciertas peculiaridades y descartar otras. Por tanto, mediante estas técnicas se abandonaron los métodos de ponderación, dejando paso a los métodos científicos. Por ejemplo, Biffen consiguió un tipo de trigo muy rico en el que, tras una larga serie de ensayos basados en las leyes de Mendel, aparecían inmunidades al moho, alta fertilidad y otras particularidades positivas.
Cuando se descubrieron los trabajos de Mendel se conocía la estructura de la célula y se observó que dentro del núcleo de cada célula hay unos cuerpos filamentosos, llamados cromosomas. El número de cromosomas en el óvulo fecundado por la combinación de dos células germinales (el caso más sencillo) es el doble, dos por clase, uno por cada progenitor. Al dividir el óculo también se divide cada cromosoma, pasando cada parte a cada célula de hijos. Así, cada nueva célula recibe un cromosoma de cada cromosoma inicial. Lo mismo ocurre con cada división siguiente. En consecuencia, cada célula vegetal y animal tiene una doble serie de cromosomas procedentes de cada progenitor.
En un principio las células germinales también tienen una doble serie de cromosomas, pero en la última transformación, es decir, la unión de los espermatozoos y los óvulos se realiza por parejas. Entonces el tipo de división es diferente: en lugar de dividir los cromosomas propiamente dichos, los de cada pareja se dividen y cada uno pasa a las células de sus hijos. Así, la célula germinal adulta recibe uno u otro de cada pareja cromosómica.
XIX. A finales del siglo XX y principios del XX, algunos expertos descubrieron la afinidad entre los efectos de la herencia de Mendel y los fenómenos celulares. Pero lo que se le ha dado formulación científica fue Sutton. Sutton observó que los cromosomas tienen una distribución como factor hereditario y que en cada caso los pares de factores y los cromosomas se distribuyen independientemente de la otra. Sin embargo, dado que el número de factores hereditarios respecto a las paredes del cromosoma es muy elevado, hay factores diferentes que están combinados en el mismo cromosoma y, por tanto, que aparezcan unidos es perfectamente normal.
A partir de 1910 Morgan y sus compañeros analizaron más a fondo estas relaciones. Se encontró una relación numérica entre el número de grupos de cualidades de la herencia y el número de pares de cromosomas. Esta cantidad es de siete para el guisante, ocho para el trigo, veinte para la rata y treinta y tres para el hombre. Por ello, aunque el número de pares de cromosomas no sea muy elevado, los tipos posibles de células germinales son muy numerosos (más de un millón) y las combinaciones posibles de dos de ellas son mucho más amplias. Por ello, es fácil entender por qué dos seres de la misma raza no se forman exactamente igual.
Mientras la investigación de la herencia iba en la línea de Mendel, también se desarrolló otro tipo de investigación, el camino de la estadística. Para ello había que tener en cuenta grandes cantidades y a los cambios humanos se adecuó la teoría de la probabilidad y, con ella, el margen estadístico de la combustión.
La curva normal de la combustión se obtiene por un número muy elevado de elementos, pero De Vries clarificó los riesgos que se pueden presentar al aplicar esta teoría en el ámbito de la herencia.
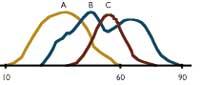
La figura superior muestra las variaciones de longitud de los tres tipos de fruta. Mientras que las longitudes se dan en línea horizontal, en vertical se marca con una longitud fija el número de elementos. Las curvas A y C son muy similares a las normales y en cada una de ellas existe una longitud media clara. Por el contrario, podemos observar que en la curva B hay al menos dos subgrupos. Si hemos tomado las tres frutas a la vez, en lugar de tener tres curvas, sólo tendríamos una, y esto tendría la apariencia de normal. Por lo tanto, tomando datos a menudo en blanco, no se puede saber si todos los elementos recibidos son del mismo tipo o de distintos subgrupos.
Algunos investigadores analizaron el comportamiento de los elementos vacíos. Partían de un único elemento para formar un grupo vacío (por ejemplo, un baberrun), y mediante la autofecundación conseguían los descendientes. Los cambios que presentaban estos elementos vacíos se ajustaban a la ley de la combustión. Pero estos cambios no pasaban por la herencia, es decir, tomando los elementos más grandes de ese grupo conseguido, sus descendientes no eran mayores, sino que se dirigían a un tamaño normal del grupo.
Al margen de la investigación de los elementos puramente enunciados, la mezcla de caracteres antiguos en cualquier raza natural da lugar a cambios que se transmiten de padres a hijos desde la antigüedad. Si los dos padres eligen para transmitir una peculiaridad, existe una mayor probabilidad de que los hijos adquieran esa peculiaridad. Por ejemplo, si tomamos altura para investigar en humanos, si los padres son largos, es decir, mayores que la media de la población, los hijos tenderán a ser así.
Pero debido a las investigaciones del Sr. Vilmorin, que en su día pasaron como las de Mendel en el lecho, los mejores resultados en el crecimiento de las plantas no se consiguen eligiendo como padres unos elementos especiales, sino seleccionando elementos de buen rendimiento medio.
Durante mucho tiempo biometristas y vaqueros tuvieron fuertes debates. Ahora se consideran complementarias las dos vías para la adecuada investigación de la herencia, por lo que deben tenerse en cuenta ambas.
Zu idazle
Zientzia aldizkaria

