

As bacterias tamén teñen esqueleto
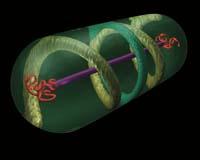
Cachorros confusos. Até hai pouco as bacterias eran nos ollos dos biólogos: sacos que conteñen material xenético, algunhas encimas e outras moléculas mesturadas; sen estruturas estables, sen orde, sen esqueletos. Pero as cousas están a cambiar. Dentro destes sacos atopáronse diversas estruturas. Moitos deles equivalentes aos compoñentes do citoesqueleto de células eucariotas.
Nas células eucariotas, é dicir, de plantas, animais, fungos e protistas, é moi coñecido o armazón interior denominado citoesqueleto. É una estrutura dinámica, composta por tres tipos de compoñentes: microfilamentos (actina), filamentos medios e microtúbulos (tubulina). E cumpre varias funcións: dar e manter forma ás células, facilitar o movemento, transporte intracelular e división celular...
Considerábase que as bacterias non tiñan citoesqueleto, polo que se consideraba que era o desenvolvido tras a formación de eucariotas (procedentes de bacterias). Pero agora parece que non foi así: a orixe das actinas, filamentos medios e tubulinas podería estar en bacterias.
As bacterias pasaron de ser protagonistas secundarios a estar de moda no campo da bioloxía celular, e os descubrimentos sucédense.
Detrás das formas

Pódese dicir que en 2001 a roda dos descubrimentos tomou o ritmo. Nese ano, un grupo de investigadores da Universidade de Newcastle en Inglaterra conseguiu por primeira vez ver a estrutura da proteína MreB dentro da bacteria Bacillus subtilis: As fibras MreB forman una hélice baixo a parede das bacterias cilíndricas.
A proteína MreB é moi parecida á actina dos eucariotas e pode ter o mesmo orixe. Ademais, cumpren una función similar. Nas bacterias cilíndricas as proteínas MreB e moi próximas a ela teñen una función estrutural, é dicir, dan forma á bacteria.
De feito, os investigadores han visto que si fáltalles esta proteína cambia o seu aspecto. B. subtilis, o MreB afecta á anchura da bacteria e o Mbl á lonxitude; e E. As bacterias coli pasan de ser cilíndricas a esféricas cando se lles quita o MreB.
De feito, as bacterias esféricas non teñen MreB. Máis aínda, analizando os xenomas de diversas bacterias e arqueos (os dous grupos de procariotas), os investigadores de Newcastle descubriron que os procariotas cilíndricos, filamentosos e helicoidales conteñen un ou máis xenes da proteína MreB, mentres que os esféricos teñen ningún.
Por outra banda, en formas máis complexas de certas bacterias é moi probable que o citoesqueleto estea relacionado. De momento, só atoparon un exemplo: As bacterias Caulobacter crescentus teñen forma de fabas, e parece que a outra proteína dálles esa curvatura. Ao retirar esta proteína estas bacterias adquiren forma cilíndrica. Esta proteína é krescentina e é moi parecida ao filamento medio dos eucariotas.
Claves da división
Á marxe das formas, o citoesqueleto tamén ten gran importancia na división celular. E niso os procariotas non teñen nada en común. As bacterias presentes no medio apropiado poden dividirse cada media hora. Pois una das claves desta eficacia pode estar precisamente no citoesqueleto. Pero os científicos non fixeron máis que empezar a aclarar os mecanismos.
A proteína FtsZ é un dos protagonistas da división celular dos procariotas. Forma un anel na zona onde se realiza a división, no centro da célula. Ao retirar esta proteína aos bacilos cilíndricos, estíranse e estiran sen dividirse en dous. Pola súa banda, os Staphylococcus aureus esféricos, una vez eliminado o xene de FtsZ, comezan a construír a nova parede celular en calquera lugar e poden chegar a aumentar o volume oito veces antes de que explote.
Dalgunha maneira, a proteína FtsZ controla onde hai que construír a nova parede. Ao seu ao redor recolle as proteínas que constrúen a parede. No caso de S. aureus forma o anel no ecuador e goberna a creación dos dous novos hemisferios. Nos bacilos, pola súa banda, parece que nun principio a proteína MreB provoca o estiramento da célula e logo entra en xogo o FtsZ, creando novos extremos redondeados paira dividir a célula en dúas.
Os aneis de FtsZ poden ter gran importancia na orixe da división celular. Un grupo de investigadores da Universidade Duke de Carolina do Norte viu recentemente que esta proteína tamén forma aneis dentro das pingas microscópicas de aceite. Agora, aínda que a división celular das bacterias é un mecanismo máis complexo, os investigadores creen que as células máis primitivas terían suficiente FtsZ paira dividirse en dous.
Por outra banda, algúns orgánulos de células eucariotas, as mitocondrias e os cloroplastos utilizan o anel de FtsZ paira dividirse. Isto reforza, en definitiva, a crenza de que estes orgánulos son procariotas que no seu día foron introducidos nunha célula eucariota.
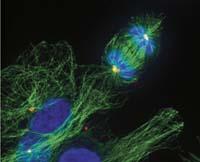
Ferramentas modificadas
Os eucariotas tamén teñen una especie de proteína FtsZ, a tubulina. E a tubulina tamén participa na división celular, pero cumpre una función moi diferente: mantén afastados os cromosomas correspondentes a cada nova célula durante a división. Desta forma asegúrase que cada nova célula terá os seus cromosomas.
É curioso que estas dúas proteínas dun mesma orixe cumpran una función tan diferente. É dicir, FtsZ e a tubulina son dúas versións dunha mesma ferramenta que os procariotas e eucariotas utilizan paira realizar traballos moi diferentes. Paira o mesmo traballo utilizan ferramentas moi diferentes. Por unha banda, paira o traballo da proteína FtsZ, equivalente á tubulina, os eucariotas utilizan una ferramenta de actina. E doutra banda, os procariotas fan o que os eucariotas fan coa tubulina cunha proteína equivalente á actina.
E é que, do mesmo xeito que os eucariotas, as bacterias tamén deben distribuír ben o material xenético na fragmentación. As bacterias posúen un único cromosoma, ademais duns aneis de ADN chamados plasmidos. Os plásmidos achegan resistencia aos antibióticos e outros 'extra' ás bacterias, polo que é moi importante que as novas bacterias queden cunha copia dos plásmidos.
Pois do mesmo xeito que fan as tubulinas cos cromosomas dos eucariotas, a proteína ParM produce a separación dalgúns plasmidos, afastando as copias. E ese ParM é una versión bacteriana da actina.
ParM é una cadea de proteínas con capacidade de autoadición e eliminación de unidades. A cadea ParM únese ás dúas novas copias do plásmido a repartir. Posteriormente, mediante a adición de novas unidades iníciase o estiramento da cadea, que envía os dous plásmidos aos dous extremos da célula.
Este é o mecanismo de distribución dalgúns plasmidos. Pero que pasa co cromosoma de bacterias? Como distribúen as copias do cromosoma? Iso segue sendo un misterio.
E non é a única. Queda un longo camiño por percorrer paira comprender ben a organización e funcionamento das células dos procariotas. Ademais dos xa mencionados, atoparon moitos máis elementos do citoesqueleto e atópanse. Tamén proteínas que non teñen nada no citoesqueleto dos eucariotas. Pero pouco sabemos dos mecanismos que están detrás de todos estes elementos. Aínda queda moito por investigar dentro destes cachorros.
Zu idazle
Zientzia aldizkaria


